 2017, Vol. 37
2017, Vol. 37



2. 浙江农林大学集贤学院, 临安 311300;
3. 金正大生态工程集团股份有限公司, 临沭 276700;
4. 浙江农林大学环境与资源学院, 临安 311300;
5. 浙江绿鑫晨农业开发有限公司, 龙游 324400;
6. 佛山科学技术学院环境与化学工程学院, 佛山 528000
2. Jixian Honors College, Zhejiang A & F University, Lin'an 311300;
3. Kingenta Ecological Engineering Group Co., LTD, Linshu 276700;
4. School of Environmental and Resource Sciences, Zhejiang A & F University, Lin'an 311300;
5. Zhejiang Lvxinchen Agricultural Development Limited, Longyou 324400;
6. School of Environment and Chemical Engineering, Foshan University, Foshan 528000
由于工业化进程的加剧、生活垃圾的处理不当及农药的滥用,土壤污染现象日益严重,其中,土壤的重金属污染较为常见.据报道,我国土壤重金属超标率达16.1%(环境保护部,2015),每年受重金属污染的粮食(如“镉米”、“砷米”等)达1000多万t,造成直接经济损失达200余亿元(周宝宣等,2015).当土壤中重金属含量超过环境自净能力并积累到一定程度后,不仅会导致土壤退化、农作物产量及品质下降,还会通过径流、淋失作用污染水源,危害人体健康(Singh et al., 2011).“十三五”规划已将土壤污染防治行动作为六方面任务之首,《土壤污染防治行动计划》(“土十条”)的出台,也体现了政府对土壤污染防治的重视.
近年来,国内外学者在土壤污染修复技术领域开展了深入的研究工作,在理论研究和工程应用上均取得了一定的成果(向捷等,2014).其中,植物修复作为一种具有广阔前景和良好效益的处理方法,备受学者关注.植物修复技术是利用植物及其根际圈微生物体系的吸收、降解等作用机制来清除环境中污染物的一项新兴污染环境治理技术(周启星等,2004).与传统修复技术相比,植物修复技术具有绿色环保、成本低廉、操作简便、对土壤的干扰小、易于为公众接受等优点(胡鹏杰等,2011).目前,利用植物修复重金属污染土壤主要是利用超积累植物富集和提取土壤中的重金属(Baker et al., 1994).然而,一般超积累植物生物量相对较小、生长缓慢、根系浅,每年从土壤中提取的重金属含量有限(Hammer et al., 2003).研究发现,利用生物量大、生长迅速、积累能力相对较强的植物也能较快地提取土壤中的重金属(Komárek et al., 2008),而速生乔木如某些柳属、杨属植物就具有这些特点,从而成为新的研究热点(Vervaeke et al., 2003;Liu et al., 2013).
竹柳(Salix sp.)为杨柳科(Salicaceae)柳属(Salix)植物,乔木,具有生长快、抗性强、材质好、杆形饱满、尖削度小等优点(张健等,2009),用途广泛,可用于工业原料林,同时也是园林绿化、农田防护林等的理想树种.近年来,竹柳先后在山东、黑龙江、上海(朱继军等,2012)、江苏、云南、安微、广西、福建、天津、湖南、贵州(胡朝慧,2014)等十多个省市引种栽培.在前期研究中,本课题组探究了竹柳对重金属的生理响应(胡国涛等,2016a),并发现竹柳可作为潜在的植物修复材料(胡国涛等,2016b).而土培条件下竹柳对重金属的富集、转移规律的研究仍处于空白阶段,竹柳在修复重金属污染土壤实际应用方面的理论尚不完善.因此,本研究采用盆栽试验,检测在重金属复合污染土壤中生长的竹柳三号及五号的生物量、光合色素、重金属含量等指标,旨在探讨不同竹柳品种对土壤中重金属的富集及转移特性,以期为竹柳作为修复重金属复合污染土壤的新型植物材料提供理论依据与数据参考.
2 材料与方法(Materials and methods) 2.1 试验材料选用竹柳三号、竹柳五号作为供试植物(浙江绿鑫晨农业开发有限公司),作为竹柳的2个不同品种,两者均有生长快、材质好等优点.据报道,竹柳三号有较高的耐盐性(朱继军等,2012).相比于竹柳三号,竹柳五号具有树皮厚、抗寒能力强的特点.挑选粗细均匀、直径为1 cm的一年生的竹柳茎段,剪成16 cm长的插条,用去离子水洗净后备用.
供试土壤共2种,分别为采自浙江省富阳市用于农业种植的水稻土(土壤Ⅰ)及该区域内某废弃的炼铜厂附近受重金属复合污染的水稻土(土壤Ⅱ),取样深度均为0~20 cm.土壤在自然条件下风干,去除肉眼可见的植物根、枯落物等,研磨,过4 mm不锈钢筛,备用.供试土壤基本理化性质如表 1所示.经测定,土壤Ⅰ中0.1 mol·L-1 CaCl2可提取态Cd含量为0.23 mg·kg-1,前期水培试验结果表明(胡国涛等,2016b),竹柳在1.12 mg·L-1的Cd处理条件下生长不受抑制,所以本试验以土壤Ⅰ作为对照.
| 表 1 供试土壤基本理化性质 Table 1 Selected physicochemical properties of the soils used |
试验在浙江农林大学智能温室(30°19′N,119°35′E)进行,自然光照,环境温度为25~33 ℃.共设土壤Ⅰ×竹柳三号、土壤Ⅰ×竹柳五号、土壤Ⅱ×竹柳三号、土壤Ⅱ×竹柳五号4个处理,每个处理5次重复.在2种供试土壤中分别加入0.15 g·kg-1 N、0.08 g·kg-1 P和0.1 g·kg-1 K,混匀后装盆,每盆3 kg干土.加入去离子水,维持田间持水量的70%,在温室中培养7 d后,每盆插入一支竹柳营养枝.日常管理主要是松土、浇水、防治病虫害,保证植物正常生长.生长100 d后,收集竹柳各部位样品,进行各项指标的测定.
2.3 测定方法收获竹柳时,用卷尺测量每盆中竹柳的株高.收获竹柳后,其根系先用自来水反复冲洗多次,再用20 mmol·L-1 EDTA浸泡15 min,以清除根系表面吸附的重金属离子,然后再用去离子水冲洗5~6次.将植物样品分为根系、枝条、叶片3部分,分别称取各部分鲜重,选取定量的新鲜竹柳叶片用于叶绿素含量的测定,剩余部分均放在烘干箱中,105 ℃下杀青30 min后,于65 ℃下烘干48 h至恒重,称重后用不锈钢粉样机将各部分分别粉碎,用于重金属含量的测定.采用丙酮乙醇混合液浸提法测定叶绿素含量(张宪政,1986), 由Arnon公式计算叶绿素含量.
竹柳生长100 d时,于上午9:00—11:00取每盆完全展开的功能叶(从茎顶端往下数第6片叶子),用OS5P调制式叶绿素荧光分析仪(北京澳作生态仪器有限公司生产)测定竹柳叶片的荧光效应.将叶夹夹在不同处理功能叶的中部,测定前叶片暗适应20 min,光化学强度为400 μmol·m2·s-1,饱和闪光强度为8000 μmol·m2·s-1.测定最大原初光能转化效率(Fv/Fm)、实际光化学量子效率Y(Ⅱ)、光化学荧光淬灭系数(qP)、非光化学荧光淬灭系数(NPQ)和电子传递速率(ETR)等指标,每项测定重复4次.以同样的方式,用便携式光合作用仪(LC-PRO+,上海旦鼎国际贸易有限公司)测定竹柳叶片的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)等指标.
分别称取过60目筛的不同部位的植物样0.3000 g,置于消煮管中,加入优级纯浓HNO3 5 mL,管口盖小漏斗(过夜)后在消煮炉(ED36,北京莱伯泰科仪器有限公司)上140~150 ℃条件下消煮至溶液呈透明无色或仅剩淡绿色(剩余约1 mL),用1%HNO3溶液定容到25 mL,过滤,滤液中重金属含量采用电感耦合等离子体发射光谱(ICP-OES,Optima 2000,PerkinElmer Co.,美国)测定.
2.4 富集系数和转移系数富集系数(BCF)和转运系数(TF)的计算方法如下(Zayed et al., 1998):
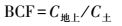
|
(1) |

|
(2) |
式中,C地上、C根、C土分别为地上部、根部和土壤重金属含量(mg·kg-1)
2.5 数据分析数据结果用Excel 2010计算,用SPSS 19.0进行分析(Analysis of variance,ANOVA),应用Origin 8.0软件作图.
3 结果(Results) 3.1 竹柳不同部位对重金属的富集状况竹柳根系、枝条、叶片中的Cd含量(均以干重计)如图 1所示.土壤Ⅱ中竹柳各部位的Cd含量显著(p<0.05) 高于土壤Ⅰ,其中,叶片中Cd含量增加最多,土壤Ⅱ中竹柳三号和竹柳五号的Cd含量分别为土壤Ⅰ的7.44和6.62倍;其次是根系,分别为土壤Ⅰ的4.54和4.37倍;再次是木质部,分别为土壤Ⅰ的1.96倍和1.80倍;最后是韧皮部,分别为土壤Ⅰ的1.94和1.93倍.各器官中的Cd含量也是叶片中最多,大小顺序为:叶片>根系>韧皮部>木质部.竹柳三号与竹柳五号相比,各部位器官中Cd含量均略高于竹柳五号, 其中,叶片中含量相差最大,为竹柳五号的1.12倍.
 |
| 图 1 竹柳各部位Cd的含量(相同系列不同小写字母表示不同处理间差异显著(p<0.05),下同) Fig. 1 Concentration of Cd in different organs of bamboo-willow plants |
由图 2可知,土壤Ⅱ中竹柳各部位中的Zn含量(均以干重计)显著(p<0.05) 高于土壤Ⅰ,其中,叶片中Zn含量增加最多,土壤Ⅱ中竹柳三号和竹柳五号叶片Zn含量分别为土壤Ⅰ的3.08和3.55倍;其次是木质部,分别为土壤Ⅰ的2.29和1.70倍;再次为根系,均为土壤Ⅰ的1.70倍;最后是韧皮部,均为土壤Ⅰ的1.23倍.各器官中的Zn含量也是叶片中最多,2种竹柳叶片中的Zn含量均超过1940 mg·kg-1.各器官中Zn含量的大小顺序为:叶片>根系>韧皮部>木质部.与竹柳五号相比,除了根系外,其余部位器官的重金属含量均是竹柳三号较高.
 |
| 图 2 竹柳各部位Zn的含量 Fig. 2 Concentration of Zn in different organs of bamboo-willow plants |
富集系数(BCF)可用于评估植物吸收和积累重金属的能力.从表 2可以看出,在重金属胁迫下,2种竹柳的木质部和韧皮部对Cd的富集系数均显著(p<0.05) 下降,叶片对Cd的富集系数显著(p<0.05) 上升.Cd在竹柳不同部位的富集系数不同,土壤Ⅰ中韧皮部富集系数最大,土壤Ⅱ中叶片的富集系数最大,富集系数最小的均为木质部.
| 表 2 不同器官对竹柳富集系数 Table 2 Bio-concentration factor of heavy metals in bamboo-willow plants |
而对于Zn来说,2种竹柳的富集系数除了叶片均有显著(p<0.05) 下降,其中,竹柳五号的木质部富集系数下降幅度远大于竹柳三号.Zn在竹柳不同部位的富集系数不同,叶片的富集系数最大,木质部最小.
3.3 不同部位对重金属的转移特性转运系数(TF)能够反映重金属在植物体内的运输和分配情况.从表 3可以看出,相较于土壤Ⅰ,土壤Ⅱ中2种竹柳的木质部与韧皮部对Cd的转移系数均有减小趋势,叶片则反之.竹柳的韧皮部与叶片对Cd都有较好的转移,TF均大于0.9.在土壤Ⅰ中,韧皮部对Cd的转移系数大于叶片,而对于土壤Ⅱ中则相反.
| 表 3 竹柳不同部位的转移系数 Table 3 Translocation factor of heavy metals in bamboo-willow plants |
对于Zn来说,2种竹柳的叶片对Zn的转移系数均有增大趋势,值得注意的是,竹柳三号木质部对Zn的转移系数显著(p<0.05) 增大,而竹柳五号木质部对Zn的转移则并没有显著(p>0.05) 影响.竹柳的韧皮部与叶片对Zn都有较强的转移能力,TF均大于0.8.相对于韧皮部,叶片的TF更大,其中,竹柳三号叶片对Zn的TF更是达到了5.04.
3.4 重金属胁迫对竹柳生物量的影响竹柳在重金属污染土壤中生长100 d后,竹柳三号及竹柳五号生长均受到不同程度的抑制.从表 4可知,与对照土壤(土壤Ⅰ)相比,重金属复合污染土壤(土壤Ⅱ)中竹柳根、茎、叶、株高显著(p<0.05) 降低,其中,竹柳三号根系、枝条、叶片的生物量分别下降了57%、76%和66%,竹柳五号的则分别下降56%、75%和62%.
| 表 4 重金属胁迫对竹柳生物量的影响 Table 4 Effect of heavy metal stress on biomass yield of bamboo-willow |
叶绿素含量的高低作为判定植物光合作用强弱的一个重要生理指标,同样是衡量植物能否作为污染土壤修复植物的指标之一(郭智等,2009).从表 5可以看出,在重金属复合污染土壤(土壤Ⅱ)中生长100 d后,与对照相比(土壤Ⅰ),竹柳的叶绿素a、叶绿素b、类胡萝卜素含量(均以鲜重计)均显著(p<0.05) 降低,而叶绿素a/b值显著(p<0.05) 升高.其中,竹柳三号的叶绿素a、叶绿素b和类胡萝卜素含量分别降低了37.5%、92%和78%,竹柳五号中3种色素含量分别降低45%、93%和83%,而2种竹柳叶片中叶绿素a/b值均升高了88%.
| 表 5 重金属胁迫对竹柳叶片中叶绿素含量的影响 Table 5 Effect of heavy metal stress on chlorophyll content in bamboo-willow leaves |
Fv/Fm是PSⅡ的最大或潜在光化学量子效率,反映PSⅡ光化学效率的高低;Y(Ⅱ)是实际光化学量子效率,反映PSⅡ反应中心处于部分开放时的实际原初光能捕获效率;NPQ是非光化学淬灭,反映天线色素吸收的光能不能用于电子传递而以热的形式耗散掉的光能部分;qP是光化学淬灭,表示PSⅡ反应中心开放部分所占的比例,代表植物光合活性高低(王威,2013);ETR是表观光电子传递效率(刘劲松等,2011),这5个指标升高均对植物生长有益.从表 6可以看出,与对照土壤相比,土壤Ⅱ中竹柳的Y(Ⅱ)和ETR显著(p<0.05) 升高,而Fv/Fm和qP差异不显著(p>0.05).对于NPQ,竹柳三号无显著(p>0.05) 差异,竹柳五号显著(p<0.05) 降低,土壤Ⅱ中的NPQ为土壤Ⅰ中的1/2.另外,在重金属复合污染土壤(土壤Ⅱ)中,竹柳三号的Y(Ⅱ)和ETR与竹柳五号的差异缩小.在对照土壤(土壤Ⅰ)中,竹柳三号的Y(Ⅱ)是竹柳五号的3.03倍,ETR是竹柳五号的3.04倍;而在重金属复合污染土壤(土壤Ⅱ)中,竹柳三号的Y(Ⅱ)和ETR则是竹柳五号的1.17倍.
| 表 6 重金属胁迫对竹柳叶片荧光效应的影响 Table 6 Effect of heavy metal stress on fluorescence effect of bamboo-willow leaves |
胞间CO2浓度(Ci)的变化方向是确定光合速率变化的主要原因是否为气孔因素的判断依据(陈根云等,2008);气孔导度(Gs)代表了植物气孔传导CO2和水汽的能力;净光合速率(Pn)是光合系统功能的直接体现,其数值升高对植物生长有益(董江超等,2015).从表 7可以看出,与对照土壤相比,土壤Ⅱ中竹柳的胞间CO2浓度显著(p<0.05) 升高,而蒸腾速率(Tr)、净光合速率显著(p<0.05) 降低,气孔导度则无显著差异.2种竹柳相比较,竹柳三号的胞间CO2浓度上升与蒸腾速率下降幅度更大,其中,蒸腾速率下降为对照组的44%.在重金属复合污染土壤(土壤Ⅱ)中,竹柳五号的净光合速率显著(p<0.05) 高于竹柳三号,其净光合速率是竹柳三号的1.87倍.
| 表 7 重金属胁迫对竹柳叶片光合特性的影响 Table 7 Effect of heavy metal stress on photosynthetic characteristics of bamboo-willow leaves |
一般情况下,在筛选重金属修复材料时,应综合考虑植物的重金属生物富集、转运及耐性能力(Zacchini et al., 2009).土壤Ⅱ中竹柳各器官中的Cd含量排序为:叶片>根系>韧皮部>木质部(图 1).竹柳叶片中Cd含量(61.4 mg·kg-1)最高,该值远大于旱柳叶片(17.27 mg·kg-1)(周疆丽,2011)、南林895叶片(12.61 mg·kg-1)与南林61-1叶片(29.23 mg·kg-1)中的Cd积累量(张春燕等,2013),说明竹柳对Cd有较强的吸收积累能力.普通植物的Zn含量为5~80 mg·kg-1(周疆丽,2011),而本试验中,叶片中的Zn含量高达1950 mg·kg-1,约是普通植物的25倍,根系、木质部和韧皮部Zn含量则相对较低,分别为409.7、117.1和358.1 mg·kg-1,含量排序为:叶片>根系>韧皮部>木质部(图 2),说明生长了100 d的竹柳吸收的Zn主要集中到叶片中.竹柳叶片中Zn的含量大于Villafranca杨树(1235 mg·kg-1)(Romeo et al., 2014)、旱柳(879.69 mg·kg-1)(周疆丽,2011)、辽胡杨(1011.34 mg·kg-1)和南林895(1081.18 mg·kg-1)(张春燕等,2013)中的Zn含量,说明相比于其他乔木,竹柳对Cd和Zn均有较强的吸收与富集能力.
在重金属土壤中生长的竹柳各部位中,叶片对Cd和Zn的富集系数和转移系数最大,木质部最小.这说明竹柳根部的Cd和Zn逐渐运输到竹柳的枝干和叶片,这与前人的发现一致(丁军露等,2015).本试验竹柳的生长周期为100 d,其韧皮部和叶片对Cd的富集系数大于3(表 2),说明竹柳根部向地上部分运输Cd能力较强.土壤Ⅱ中叶片对Cd的转移系数大于韧皮部,这与土壤Ⅰ中的结果相反(表 3),这是竹柳叶片对Cd的转移系数增加,而韧皮部对Cd的转移系数减少所致,说明竹柳在重金属污染土壤中生长会加强其将Cd向叶片转移的能力.叶片对Cd的转移系数大于1,远大于超富集植物长柔毛委陵菜(0.38) 的转移系数(胡鹏杰,2010),说明竹柳对Cd具有较强转移能力.而对于土壤Ⅱ中生长的竹柳对Zn的富集系数,除了叶片外,其他部位均有显著下降到低于1的程度(表 2),但其根系的富集系数(0.23) 仍大于旱10柳树(0.12) 和旱34柳树(0.16) 的富集系数(施翔等,2010),说明竹柳根系有较强的富集能力;而土壤Ⅱ处理下,叶片中Zn的富集系数仍大于1,高于柳树(0.8)(Vervaeke et al., 2003),说明竹柳在重金属土壤中,其叶片对Zn也有较强的富集能力.竹柳叶片对Zn的转移系数有增大趋势(表 3),这与汤叶涛等(2009)研究结果一致,他们发现超富集植物滇苦菜在Zn含量越高的土壤中,其地上部分转移系数越大.另外,竹柳三号木质部对Zn的转移系数有增大趋势,竹柳五号则没有明显变化(表 3).由此可以看出,竹柳三号比竹柳五号对Zn的转运能力更强.土壤Ⅱ中竹柳叶片对Zn的转移系数大于或接近于5,说明重金属胁迫下竹柳叶片对Zn的转移能力较强.
4.2 重金属胁迫下的竹柳生理响应在重金属土壤(土壤Ⅱ)中生长的竹柳,各部位器官的生物量与对照组(土壤Ⅰ)相比均显著降低,这可能是因为土壤中过量的重金属元素抑制了植物生长.Cd是植物生长生物非必须元素且极易被植物吸收并积累,过量的Cd不仅会影响作物的产量和品质,而且还会对植物和动物产生很强的毒性(申强等,2015).土壤中Zn含量达到70~400 mg·kg-1时(Kabata Pendias et al., 1984)会对植物产生毒害作用,本试验供试土壤中Zn含量明显超过这个界限,为标准的4.5倍.在Cd和Zn胁迫作用下,竹柳的生长受到了一定抑制,但其根系生物量下降量小于群众杨(58%)和中富1号柳树(63%)(张春燕等,2013),具有作为耐性植物修复土壤的潜质.
叶绿素是植物进行光合作用利用的主要色素,其含量的高低能够反映光合作用的强弱(范爱君等,2006).在复合重金属污染的土壤中(土壤Ⅱ),竹柳叶片的光合色素叶绿素a、叶绿素b、类胡萝卜素含量与对照土壤(土壤Ⅰ)相比显著降低,而叶绿素a/b却显著升高(表 5).一般情况下,叶绿素a/b的值越大,类囊体的垛叠程度越大,光抑制作用越弱,植物对光能的利用效率越高(Wang et al., 2012).本试验结果说明在重金属胁迫下,竹柳的叶绿素合成虽然受到了抑制,但叶绿素a下降的幅度要远小于叶绿素b,从而使叶绿素a/b升高,一定程度上减弱了光抑制作用,提高了光能利用率.这与含羞草、白雪姬和树马齿苋在Cd污染土壤中生长的结论类似(燕傲蕾等,2010).
在重金属胁迫下,竹柳的Fv/Fm和qP无显著变化,说明重金属胁迫对竹柳光合活性影响较小;而Y(Ⅱ)和ETR则显著上升(表 6),说明在重金属的影响下,竹柳的PSⅡ光系统的实际光化学量子效率和表观电子传递效率提高了,光能转化效率提高,为暗反应的光合碳同化积累更多所需的能量(吴家森等,2008),这与超富集植物滇苦菜在较低浓度的Cd下荧光效应变化一致(应蓉蓉等,2010).
胞间CO2浓度、蒸腾速率、气孔导度、净光合速率是反应光合作用强度的指标,是衡量植物对逆境光合生理响应能力的重要参数(周续莲等,2011).在重金属胁迫下生长100 d的竹柳,净光合速率均较对照显著下降,主要因为气孔限制和非气孔限制(Farquhar et al., 1982).Cd胁迫会对植物气孔作用产生影响(Vitoria et al., 2004),本实验并未出现该现象,这可能是因为土壤Ⅱ中的Cd并未达到竹柳气孔关闭的限值,说明竹柳对重金属Cd的耐受性较强.相比之下,竹柳五号的净光合速率较大,是竹柳三号的1.87倍,说明在重金属土壤中竹柳五号生长状况较竹柳三号好.
综上所述,2种竹柳相比,竹柳三号对Zn的转运能力大于竹柳五号,而竹柳五号的净光合速率更大.此外,土壤污染程度是竹柳重金属含量、重金属富集系数、转移系数及生理指标的主要影响因素,而竹柳品种和土壤污染程度在对竹柳荧光效应和光合特性的影响上有交互作用.
5 结论(Conclusions)竹柳在重金属严重污染的土壤中能存活生长,但其生物量、叶绿素、荧光效应、光合特性等生理指标均受到不同程度的影响.竹柳在重金属胁迫条件下生长100 d时,各器官重金属含量和转移情况不同,叶片中Cd和Zn的含量和转移系数均为最高.2种竹柳叶片对Cd的转移系数大于1,对Zn的转移系数大于或接近于5.因此,竹柳三号和竹柳五号2个竹柳品系用于重金属植物修复均有较理想的效果,在实际应用时可通过集中处理叶片,达到降低重金属二次污染的目的.
Baker A J M, Reeves R D, Hajar A S M. 1994. Heavy metal accumulation and tolerance in British populations of the metallophyte Thlaspi caerulescens J. & C.Presl (Brassicaceae)[J]. New Phytologist, 127(1): 61–68.
DOI:10.1111/nph.1994.127.issue-1
|
陈根云, 陈娟, 许大全. 2010. 关于净光合速率和胞间CO2浓度关系的思考[J]. 植物生理学报, 2010, 46(1): 64–66.
|
丁军露. 2015. 竹柳生长培育及对重金属污染土壤生态修复的相关技术研究[D]. 南京: 东南大学
http://cdmd.cnki.com.cn/Article/CDMD-10286-1016753064.htm |
董江超, 路斌, 缴丽莉, 等. 2015. NaCl胁迫对竹柳光合荧光特性的影响[J]. 河北林果研究, 2015, 30(3): 222–226.
|
Farquhar G D, Caemmerer S V.1982.Modelling of Photosynthetic Response to Environmental Conditions//Physiological Plant Ecology Ⅱ[M].Berlin Heidelberg:Springer.549-587
|
范君华, 刘明. 2006. 不同叶型零式果枝海岛棉叶片光合色素特性比较[J]. 中国棉花, 2006, 33(4): 11–12.
|
郭智, 原海燕, 奥岩松. 2009. 镉胁迫对龙葵幼苗光合特性和营养元素吸收的影响[J]. 生态环境学报, 2009, 18(3): 824–829.
|
Hammer D, Keller C. 2003. Phytoextraction of Cd and Zn with Thlaspi caerulescens in field trials[J]. Soil Use & Management, 19(2): 144–149.
|
胡朝慧. 2014. 竹柳山地造林综合技术初探[J]. 农技服务, 2014, 31(6): 160–161.
|
胡国涛, 杨兴, 陈小米, 等. 2016a. 速生树种竹柳对重金属胁迫的生理响应[J]. 环境科学学报, 2016a, 36(10): 3870–3875.
|
胡国涛, 于阳, 杨兴, 等. 2016b. 速生树种竹柳对镉的吸收、积累与分布特性[J]. 环境科学学报, 2016b, 36(4): 1508–1514.
|
胡鹏杰. 2010. 超富集植物长柔毛委陵菜和滇苦菜对镉的吸收、转运及分布机制研究[D]. 广州: 中山大学
|
胡鹏杰, 吴龙华, 骆永明. 2011. 重金属污染土壤及场地的植物修复技术发展与应用[J]. 环境监测管理与技术, 2011, 23(3): 39–42.
|
Kabata Pendias A, Pendias H K. 1984. Trace Elements in Soils and Plants[M]. Florida: CRC Press: 951–974.
|
Komárek M, Tlustoš P, Száková J, et al. 2008. The use of poplar during a two-year induced phytoextraction of metals from contaminated agricultural soils[J]. Environmental Pollution, 151(1): 27–38.
DOI:10.1016/j.envpol.2007.03.010
|
Liu W T, Ni J C, Zhou Q X. 2013. Uptake of heavy metals by trees:prospects for phytoremediation[J]. Materials Science Forum, 743-744: 768–781.
DOI:10.4028/www.scientific.net/MSF.743-744
|
刘劲松, 石辉, 李秧秧. 2011. 镉胁迫对黄瓜幼苗光合和叶绿素荧光特性的影响[J]. 水土保持研究, 2011, 18(5): 187–190.
|
Romeo S, Francini A, Ariani A, et al. 2014. Phytoremediation of Zn:Identify the diverging resistance, uptake and biomass production behaviours of poplar clones under high zinc stress[J]. Water, Air, & Soil Pollution, 225(1): 1–12.
|
申强, 吴军, 周军, 等. 2015. 植物对重金属镉(Cd)抗性的研究综述[J]. 上海农业科技, 2015(2): 30–33.
|
施翔, 陈益泰, 吴天林, 等. 2010. 7个柳树无性系在Cu/Zn污染土壤中的生长及对Cu/Zn的吸收[J]. 中国环境科学, 2010, 30(12): 1683–1689.
|
Singh B R, Gupta S K, Azaizeh H, et al. 2011. Safety of food crops on land contaminated with trace elements[J]. Journal of the Science of Food and Agriculture, 91(8): 1349–1366.
DOI:10.1002/jsfa.v91.8
|
汤叶涛, 吴妤都, 仇荣亮, 等. 2009. 滇苦菜(Picris divaricata Vant.)对锌的吸收和富集特性[J]. 生态学报, 2009, 29(4): 1823–1831.
|
Vervaeke P, Luyssaert S, Mertens J, et al. 2003. Phytoremediation prospects of willow stands on contaminated sediment:a field trial[J]. Environmental Pollution, 126(2): 275–282.
DOI:10.1016/S0269-7491(03)00189-1
|
Vitoria A P, Rodriguez A P M, Cunha M, et al. 2003. Structural changes in radish seedlings exposed to cadmium[J]. Biologia Plantarum, 47(4): 561–568.
|
Wang Q, Zhang W, Li C, et al. 2012. Phytoremediation of atrazine by three emergent hydrophytes in a hydroponic system[J]. Water Science & Technology A Journal of the International Association on Water Pollution Research, 66(6): 1282–1288.
|
王威. 2013. 蝴蝶兰不同品种叶绿素荧光特性研究[D]. 福州: 福建农林大学
http://cdmd.cnki.com.cn/Article/CDMD-10389-1013327212.htm |
吴家森, 宋福强, 陈荣, 等. 2008. 3种七叶树属植物叶片气体交换特征和叶绿素荧光特性比较[J]. 植物研究, 2008, 28(4): 438–441.
|
向捷, 陈永华, 向敏, 等. 2014. 土壤重金属污染修复技术比较研究[J]. 安徽农业科学, 2014, 42(22): 7367–7369.
DOI:10.3969/j.issn.0517-6611.2014.22.027 |
Xia H, Shu W. 2001. Resistance to and uptoke of heavy metals by Vetiveria zizanioides and Paspalum notatum from lead/zinc mine tailings[J]. Acta Ecologica Sinica, 21(7): 1121–1129.
|
燕傲蕾, 吴亭亭, 王友保, 等. 2010. 三种观赏植物对重金属镉的耐性与积累特性[J]. 生态学报, 2010, 30(9): 2491–2498.
|
应蓉蓉, 汤叶涛, 汤璐, 等. 2010. 镉对超富集植物滇苦菜(Picris divaricata Vant. )光合及叶绿素荧光特性的影响[EB/OL]. 北京: 中国科技论文在线. 2010-07-21, http://www.paper.edu.cn/releasepaper/content/201007-408
http://www.cnki.com.cn/Article/CJFDTOTAL-STXB201002007.htm |
Zacchini M, Pietrini F, Mugnozza G S, et al. 2009. Metal tolerance, accumulation and translocation in poplar and willow clones treated with cadmium in hydroponics[J]. Water, Air, & Soil Pollution, 197(1): 23–34.
|
Zayed A, Lytle C M, Terry N. 1998. Accumulation and volatilization of different chemical species of selenium by plants[J]. Planta, 206(2): 284–292.
DOI:10.1007/s004250050402
|
张健, 李玉娟, 张树清, 等. 2009. 美国竹柳生物特性与繁殖试验简报[J]. 上海农业科技, 2009(6): 116–116.
|
张春燕, 王瑞刚, 范稚莲, 等. 2013. 杨树和柳树富集Cd、Zn、Pb的品种差异性[J]. 农业环境科学学报, 2013, 32(3): 530–538.
|
张宪政. 1986. 植物叶绿素含量测定—丙酮乙醇混合液法[J]. 辽宁农业科学, 1986, 3: 28–30.
|
周宝宣, 袁琦. 2015. 土壤重金属检测技术研究现状及发展趋势[J]. 应用化工, 2015, 44(1): 131–138.
|
周疆丽. 2011. 旱柳对重金属的耐性研究[D]. 合肥: 安徽农业大学
http://cdmd.cnki.com.cn/Article/CDMD-10364-1013162966.htm |
周启星. 2004. 污染土壤修复原理与方法[M]. 北京: 科学出版社.
|
周续莲, 吴宏亮, 康建宏, 等. 2011. 不同灌水处理对春小麦水分利用率和光合速率的影响[J]. 农业科学研究, 2011, 32(4): 1–5.
|
朱继军, 陈必胜, 王玉勤, 等. 2012. 苏柳、竹柳等速生能源柳引种研究[J]. 安徽农业科学, 2012, 40(29): 14290–14291.
DOI:10.3969/j.issn.0517-6611.2012.29.053 |