 2017, Vol. 37
2017, Vol. 37



2. 福建师范大学亚热带湿地研究中心, 福州 350007;
3. 湿润亚热带生态-地理过程教育部重点实验室, 福州 350007
2. Reserach Centre of Wetlands in Subtropical Region, Fujian Normal University, Fuzhou 350007;
3. Key Laboratory of Humid Sub-tropical Eco-geographical Process of Ministry of Education, Fuzhou 350007
滨海湿地作为全球生产能力最高的生态系统之一, 在全球碳平衡和气候变化中发挥着重要作用(Tian et al., 2010).围垦养殖作为人类开发利用滨海潮滩湿地的主要方式之一, 可在一定程度上缓解沿海地区人多地少的矛盾(陈军冰等, 2012), 但围垦养殖管理方式下, 沉积环境会发生很大变化, 且人为干扰作用加大, 对湿地沉积物有机碳循环具有深刻的影响.根据沉积物有机碳库对外界因素的敏感性和周转速度, 可分为活性有机碳库和惰性有机碳库, 有机碳短期内的动态变化主要发生在易分解矿化的活性碳部分, 活性有机碳组分较为复杂, 对环境变化的响应更为敏感, 其中, 微生物生物量碳(MBC)、易氧化有机碳(EOC)和水溶性有机碳(WSOC)是其重要的表征指标(朱丽琴等, 2017).MBC主要是土壤中微生物、细菌和真菌体内所含的有机碳, 反映土壤微生物活性, 与有机碳的转化密切相关, 是最活跃的有机碳组分之一(朱丽琴等, 2017).EOC是土壤中周转最快的有机碳组分, 常作为有机碳动态变化的敏感性指标(柯跃进等, 2014).而WSOC主要来源于表层有机质的淋溶和分解, 是微生物直接利用的能源, 移动快、易溶于水(Jagadamma et al., 2014; 朱丽琴等, 2017).虽然土壤中有机碳组分所占比例小, 但这些组分对环境变化更加敏感, 与有机碳的固持和释放密切相关(Núñez et al., 2001).此外, 有机碳组分作为有机碳分解矿化的主要动力, 其化学组成和存在状态是影响有机碳矿化的关键因素(Núñez et al., 2001), 有机碳矿化作为碳循环的关键过程, 其动态变化直接影响沉积物碳元素的释放与温室气体的形成(李忠佩等, 2004).因此, 研究围垦养殖对有机碳组分及其矿化特征的影响, 对于合理开发湿地和认识湿地沉积物碳平衡具有重要意义.
目前, 越来越多的学者重视围垦过程及围垦年限对滨海湿地生态系统的影响, 且对此也已经开展了一些相关研究.例如, 丁晨曦等(2013)研究发现, 黄河三角洲盐碱地开发成人工林(刺槐)、人工耕作农田(棉花)后, 表层(0~20 cm)土壤有机质、全氮质量分数增加;同时, 也有研究指出, 随着围垦时间的延长, 土壤有机质、全氮和全磷含量均呈增加的趋势(吴明等, 2008).然而, 也有学者研究发现, 天然湿地转为草地和水稻田等用地后, 土壤SOC和EOC含量均有不同程度降低(曾从盛等, 2008).当前, 对水产养殖塘也开展了一些研究, 但主要集中在沉积物有机碳、氮、磷、硅和重金属等元素(吴明等, 2008; 曾从盛等, 2008; 吴建兰等, 2012), 而对养殖塘沉积物有机碳组分及有机碳矿化特征的研究还鲜见报道, 且以往对于养殖区沉积物的研究主要以箱网养殖和筏式养殖方式为主, 对围垦养殖方式的研究还相对较少.围垦养殖后, 形成相对封闭的水淹环境, 且人为活动(饵料添加和曝气等)对沉积物有机碳组分及其矿化分解可能具有很大影响, 因此, 很有必要对围垦养殖塘沉积物有机碳组分及其矿化特征开展研究.
基于此, 本研究以闽江河口鳝鱼滩湿地为研究区, 选取不同围垦年限(3 a和15 a)的养殖塘及短叶茳芏(Cyperus malaccensis)(养殖区围垦前主要土著植被)湿地沉积物为研究对象, 探讨围垦养殖对沉积物TOC、有机碳组分和矿化的影响, 旨在了解滨海湿地围垦后沉积物有机碳库特征的变化, 这有利于深入认识人类活动对湿地碳循环的影响, 为合理开发和保护湿地与可持续发展提供科学依据.
2 材料与方法(Materials and methods) 2.1 研究区概况闽江口自然保护区鳝鱼滩湿地(26°00′36″~26°03′42″N, 119°34′12″~119°41′40″E), 位于福建省长乐市闽江入海口, 区内属亚热带海洋性季风气候, 气候温暖湿润, 年平均气温为19.6 ℃, 年降水量为1346 mm(刘剑秋等, 2006).土壤以滨海砂质土为主, 潮汐属典型的半日潮.本研究样地布设在鳝鱼滩湿地观鸟台附近堤坝内围垦养虾塘和堤坝外短叶茳芏湿地(图 1), 鳝鱼滩湿地植物群落主要以芦苇(Phragmites australis)和短叶茳芏为主, 2002年以来, 互花米草(Spartina alterniflora)迅速入侵, 目前已成为研究区内的主要物种之一.根据调查访问, 围垦区围垦养殖前主要土著植被为短叶茳芏, 养殖塘养殖期间平均水深约1 m, 一般从6月开始放养虾苗, 11月下旬收获, 期间每天施加3次饵料, 强度约为0.23 kg· m2· d-1(杨平等, 2013).基于遥感影像和实际调查访问, 样地有2个不同年限的围垦养殖区, 其中, 南部区域在1998年开始围垦养殖, 围垦年限约为15 a, 北部区域2011年才开始围垦, 围垦年限约为3 a.
 |
| 图 1 研究区与样地布设示意图 Fig. 1 Map of study region and sampling sites |
于2014年11月11日, 在闽江口鳝鱼滩湿地选取2个不同围垦年限(3 a和15 a)的养殖塘, 从养殖塘中心向四周随机选取4个样点, 通过养殖木船, 利用深水底泥采集器, 分别采集0~10 cm和10~20 cm深度沉积物;此外, 在堤坝外短叶茳芏湿地选取4个样点, 利用不锈钢取土器, 分别采集0~10 cm和10~20 cm深度沉积物, 并用环刀法, 测定沉积物容重.样品采集后, 立即装入自封袋, 放进保温箱带回室内, 一部分放入冰箱4 ℃保鲜, 另一部分冷冻风干.风干的沉积物样品过2 mm筛, 之后再取一小部分过0.149 mm筛, 待测.
2.2.2 沉积物有机碳矿化的测定采用室内密闭恒温培养、气相色谱法相结合测定有机碳矿化速率(王莲阁等, 2014; 胡敏杰等, 2016).具体为:称取风干沉积物20.0 g, 将沉积物样品均匀铺于120 mL培养瓶, 用蒸馏水调节沉积物湿度至饱和持水量的60%;密闭培养前, 将培养瓶放入恒温培养箱(17.4 ℃)内预培养1周, 以恢复沉积物中微生物活性; 再用704硅胶密封培养瓶和塞子之间的空隙(塞子的中间有2个小孔, 内插玻璃管, 管外再套一段硅胶软管, 以适合的硅胶塞塞紧软管通口, 作为气体采样口);将装置好的培养瓶放入17.4 ℃恒温培养箱中培养56 d, 正式培养后, 分别测定第1、3、5、7、10、14、21、28、38和56 d的有机碳矿化速率;在每个采样日采集气体前, 用高纯度N2对每个培养瓶进行冲洗(约3 min)后密封, 用气密针从培养瓶内抽取10 mL气体, 作为测定初始值并及时补充10 mL高纯度N2, 在24 h后, 再次抽10 mL气体并补充同体积高纯度N2, 气体采集结束后, 及时用气相色谱仪测定CO2浓度, 根据两次的浓度计算该培养期的有机碳矿化速率;培养期间每周采用称重法保持沉积物含水量的恒定.
2.2.3 沉积物有机碳组分及基本理化性质测定沉积物TOC测定采用重铬酸钾-外加热法;MBC测定采用氯仿熏蒸-0.5 mol· L-1硫酸钾浸提法, 浸提液中有机碳浓度用TOC-VCPH分析仪测定, 并根据MC=EC/0.38(EC为熏蒸与未熏蒸沉积物有机碳含量差值)公式计算MBC含量MC (鲁如坤, 2000);EOC含量采用333 mmol· L-1 KMnO4氧化比色法测定(鲁如坤, 2000);采用去离子水浸提法测定WSOC含量(高灯州等, 2016; 胡敏杰等, 2016), 具体为:称取10 g过2 mm筛的新鲜沉积物于100 mL离心管中, 按沉积物与水1:5的比例加入50 mL去离子水, 在25 ℃恒温振荡器以250 r· min-1的速度振荡30 min, 再以10000 r· min-1的转速离心20 min;离心后的上清液用去碳处理过的0.45 μm的滤膜过滤, 采用TOC-VCPH仪器测定浸提液有机碳含量, 计算沉积物WSOC含量.采用奥立龙868型酸度计和DDS-307型电导仪测定沉积物酸碱度(pH)和电导率(EC);容重采用环刀法测定, 采用烘干法测定沉积物含水量(鲁如坤, 2000);采用邻菲罗啉比色法测定Fe2+和Fe3+含量, 并计算Fe2+/Fe3+(Luo et al., 2014);用土壤碳氮元素分析仪(Elementar Vario MAX CN, Germany)测定沉积物全氮(TN)含量;浓硫酸-高氯酸消煮, 钼锑抗比色法测定全磷(TP)含量(Ruban et al., 2001);粒径采用激光粒度分析仪(Master Sizer 2000, UK)测定, 并且根据国际制土壤质地分类标准分为黏粒(<2 μm)、粉粒(2~20 μm)和砂粒(>20 μm)(李顺姬等, 2010).供试沉积物基本理化性质如表 1所示.
| 表 1 供试沉积物基本理化性质 Table 1 Basic physical and chemical properties of tested sediment samples |
采用SPSS19.0软件对数据进行方差分析和显著性检验, 其中, 显著性差异p<0.05.采用Origin 8.0作图.
本研究采用有机碳矿化速率和累积矿化量两个指标作为有机碳矿化差异的评价指标, 其中, 有机碳(CO2-C)矿化速率计算公式见式(1).
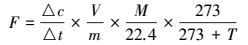
|
(1) |
式中, F为CO2-C矿化速率(μg· g-1· d-1), Δc/Δt为培养瓶内单位时间内(24 h)浓度的变化量(μg· d-1· g-1), V为培养瓶体积(L), m为沉积物干重(g), M/22.4是标准气体质量密度(g· L-1), T是培养温度(℃).有机碳(CO2-C)累积矿化量则为培养时间内有机碳矿化速率的累加(μg·g-1).此外, 由于培养时间为56 d, 有机碳矿化主要为活性有机碳分解, 因此, 采用一级动力学方程(式(2))模拟分析不同处理下有机碳矿化动态(Cooper et al., 2011).
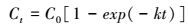
|
(2) |
式中, Ct为t时刻有机碳累积矿化量(mg· g-1), C0为有机碳的潜在矿化量(mg· g-1), t为培养天数(d), k为矿化速率常数(d-1).
3 结果与分析(Results and analysis) 3.1 围垦养殖塘与短叶茳芏湿地沉积物TOC及有机碳组分含量闽江口围垦养殖塘与短叶茳芏湿地TOC及有机碳组分含量存在一定的差异(图 2).围垦养殖3 a后的沉积物表层(0~10 cm)TOC含量无显著变化, 但围垦养殖15 a后的沉积物表层TOC含量显著增加(p<0.05), 达到(29.89 ±1.76) g· kg-1, 显著大于短叶茳芏湿地((17.75±0.89) g· kg-1)和围垦3 a((20.07±2.35) g· kg-1)沉积物, 而亚表层(10~20 cm)养殖塘与短叶茳芏湿地沉积物TOC含量无显著差异(p>0.05)(图 2a).短叶茳芏和养殖塘沉积物MBC含量为98.88~192.81 mg· kg-1, 不同沉积层养殖塘和短叶茳芏湿地MBC含量均无显著差异(p>0.05)(图 2b).围垦后, 沉积物EOC含量有所降低, 其表层沉积物EOC含量表现为短叶茳芏湿地和围垦3 a养殖塘高于围垦15 a养殖塘, 而亚表层表现为短叶茳芏湿地>围垦15 a养殖塘(p<0.05)(图 2c).围垦养殖后沉积物WSOC分布特征与EOC相反, 其中在0~10 cm深度, 围垦15 a养殖塘沉积物WSOC含量为(377.87±37.15) mg· kg-1, 显著高于围垦3 a养殖塘((270.72±23.32) mg· kg-1)和短叶茳芏湿地((224.91±30.56) mg· kg-1), 围垦3 a养殖塘和短叶茳芏湿地沉积物WSOC含量无显著差异, 而在10~20 cm深度, 围垦3 a和15 a养殖塘的WSOC含量均显著高于短叶茳芏湿地(p<0.05)(图 2d).
 |
| 图 2 短叶茳芏湿地与养殖塘沉积物有机碳组分分布特征 (同一土层深度不同字母表示数据间差异显著, p<0.05) Fig. 2 Distribution characteristics of organic carbon fractions in sediments of C. malaccensis wetland and aquaculture ponds |
闽江河口养殖塘与短叶茳芏湿地沉积物有机碳矿化速率如图 3所示.为期56 d的培养期间, 养殖塘与短叶茳芏湿地有机碳矿化速率在培养初期(第1~7 d)较大, 并在第3 d达到峰值; 培养后期(7~56 d), 有机碳矿化速率则随着培养时间延长呈下降趋势并趋于平稳.整个培养期间, 养殖塘表层沉积物有机碳矿化速率均显著高于短叶茳芏湿地(p<0.05), 表现为围垦15 a养殖塘>围垦3 a养殖塘>短叶茳芏湿地, 而亚表层各类型沉积物有机碳矿化速率总体差异不显著(p>0.05).此外, 培养前期表层有机碳矿化速率略高于亚表层, 而培养后期差异不显著(p>0.05).
 |
| 图 3 短叶茳芏湿地与围垦养殖塘沉积物CO2-C矿化速率变化特征 Fig. 3 Changes of CO2-C mineralization rate of sediments in C. malaccensis wetland and aquaculture ponds |
在培养初期, 由于可利用性碳源较多, 有机碳累积矿化量增长较快, 之后逐渐减慢趋于平稳(图 4).为期56 d的培养后养殖塘累积矿化量显著高于短叶茳芏湿地(p<0.05), 其中, 表层有机碳累积矿化量大致表现为围垦15 a养殖塘(2178 μg· g-1)>围垦3 a养殖塘(1280 μg· g-1)>短叶茳芏湿地(908 μg· g-1), 而亚表层, 则表现为围垦3 a养殖塘(1086 μg· g-1)>围垦15 a养殖塘(1035 μg· g-1)>短叶茳芏湿地(772 μg· g-1).
 |
| 图 4 短叶茳芏湿地与围垦养殖塘沉积物CO2-C累积矿化量变化特征 Fig. 4 Changes of CO2-C cumulative mineralization of sediments in C. malaccensis wetland and aquaculture ponds |
围垦养殖塘和短叶茳芏湿地不同深度沉积物有机碳矿化累积动态都符合一级动力学指数方程, 均达到较好的效果(R2>0.964, 表 2).由表 2可以看出, 不同类型沉积物C0存在较大差异, 变化范围为741~2068 μg· g-1, 其中围垦养殖塘高于短叶茳芏湿地, 且表层沉积物C0略高于亚表层.k值正好与C0相反, 相同深度k值随C0增加而减小.进一步计算C0/TOC发现, 同一沉积层, 养殖塘C0/TOC比值均高于短叶茳芏湿地.
| 表 2 沉积物有机碳矿化的一级动力学参数及C0/TOC值 Table 2 Parameters of the first order kinetics and C0/TOC values for the sediment of organic carbon mineralization |
有机碳组分、理化性质和有机碳矿化指标之间的相关分析显示(表 3), TOC、MBC和WSOC三者之间具有显著相关关系(p<0.05), 而与EOC均无显著相关关系.TOC与TN、TP和黏粒含量呈显著正相关(p<0.01), WSOC与Fe2+/Fe3+、TN和TP呈显著正相关(p<0.05), 而MBC和EOC与理化性质均无明显相关性.累积矿化量、C0与TOC、WSOC、Fe2+/Fe3+、TN、TP和黏粒呈显著相关关系(p<0.01).
| 表 3 有机碳组分、理化性质与有机碳累积矿化指标之间的相关系数 Table 3 Relationship between organic carbon fractions, properties and organic carbon cumulative mineralization indexes |
土地利用方式的转变会直接影响土壤有机碳含量和组成, 进而改变有机碳的分解与转化(佘建等, 2014).本研究表明, 围垦养殖总体上增加了表层TOC, 尤其是围垦15 a的养殖塘TOC增加显著, 这一方面是因为养殖塘长期处于淹水环境中, 厌氧的环境抑制了有机质的分解, 而短叶茳芏湿地干湿交替的环境相对易于有机质分解(张静等, 2014).另外, 养殖期间投放的饵料中富含大量有机质可能是其沉积物有机碳含量增加的另一重要原因.有学者基于养虾池中的营养物质平衡研究表明, 在养殖过程中只有约6%的有机质被吸收利用, 其它多以各种形式进入沉积物-水界面环境中, 且很大部分沉积下来(Funge-Smith et al., 1998);同时也有研究指出, 人为添加的饵料虽然大部分被鱼虾所摄食, 但约有30%未被摄食, 这些残饵和生物体排泄物等经过分解后, 主要蓄积在表层沉积物(陈宏友等, 2004), 这可能是造成养殖塘表层沉积物TOC含量显著增加的主要原因.再者, TOC含量还受沉积物粒径、TN和TP等理化性质的影响(王绍强等, 2008; 李顺姬等, 2010), 已有研究表明, 黏粒可以吸附有机质, 提高有机碳含量(李顺姬等, 2010), 而碳、氮和磷作为生态系统中最主要的生源要素, 三者之间有密切的相互耦合关系(王绍强等, 2008).本研究显示的TOC与黏粒、TN和TP显著正相关关系也进一步证实这一观点.此外, 与围垦15 a的养殖塘相比, 围垦3 a的养殖塘表层沉积物TOC含量增加不显著, 这可能是因为随着围垦年限的增加, 表层有机物质具有一定的富集现象, 围垦3 a的养殖塘围垦时间相对较短, 有机物的累积还相对较少, 故TOC含量增加不显著.
有机碳组分具有易矿化分解、水溶性强和周转时间短等特点, 相比TOC, 有机碳组分对环境变化的响应更加敏感(Laik et al., 2009).MBC作为最活跃的有机碳组分之一, 可在一定程度上表征微生物活性的强弱, 与土壤有机碳的转化密切相关(朱丽琴等, 2017).本研究显示, 围垦养殖后MBC含量无显著变化, 理论上, 养殖塘饵料等有机质的输入, 为微生物利用提供能量, 其沉积物MBC含量会有所增加, 本研究出现结果不一致的原因可能与沉积环境有关.养殖塘长期水淹和饵料分解消耗大量的氧气, 使养殖塘形成高度厌氧环境, 微生物的活性受到一定的影响, 故很难维持较高的微生物量(张静等, 2014), 使得MBC含量增加不显著.由本研究Fe2+/Fe3+比例可知(表 1), 围垦养殖后Fe2+/Fe3+显著升高, 说明围垦养殖后, 沉积物还原环境增强, 这也进一步证实了上述假设.EOC是土壤中移动快、易氧化的有机碳组分(柯跃进等, 2014).本研究显示养殖塘EOC含量有所下降, 尤其是围垦15 a的养殖塘, 其原因主要是养殖塘相对厌氧的环境致使有机碳中易氧化部分含量降低, 这也与曾从盛等(2008)的研究结果相一致.WSOC是表层有机质淋溶和微生物分解后形成的代谢产物, 是微生物生长的能量来源, 易溶于水(Jagadamma et al., 2014; 柳敏等, 2007).本研究表明, 围垦养殖后沉积物WSOC含量总体上显著增加, 这是因为短叶茳芏湿地受潮汐作用的影响, 潮汐作用下, 土壤中的WSOC容易流失(Jagadamma et al., 2014), 高灯州等(2016)通过原位培养实验显示, 高潮滩土壤-植物系统搬至中潮滩后, 土壤WSOC含量显著降低, 本研究的养殖塘较为封闭且处于稳定的水淹环境, 这可能使得养殖塘与短叶茳芏湿地沉积物MBC含量无显著差异的情况下, 养殖塘沉积物SWOC含量显著高于短叶茳芏湿地的重要原因.
4.2 围垦养殖对湿地沉积物有机碳矿化的影响有机碳矿化是指微生物直接利用土壤中的生源要素(C、N、P等)来完成自身代谢, 分解有机质的同时释放CO2(张林海等, 2011).本研究显示, 养殖塘沉积物有机碳矿化速率和累积矿化量显著高于短叶茳芏湿地沉积物, 这表明围垦养殖促进了沉积物有机碳矿化, 这可能与沉积环境、沉积物养分含量及其理化性质等因素变化有关.相关研究(李顺姬等, 2010)指出, 沉积物养分含量是影响有机碳矿化的主要因素, 养分越充足, 微生物活性越高, 其矿化越强.本文研究结果显示, 围垦养殖后, 沉积物TN、TP和TOC总体上显著增加(表 1和图 2a), 这可能是导致养殖塘沉积物有机碳矿化速率和累积矿化量增加的原因之一.相关分析结果显示, 沉积物有机碳累积矿化量和潜在矿化量(C0)与TN、TP和TOC均呈显著正相关(表 3), 这进一步证实了上述结果.另外, 研究表明有机碳组分与有机碳的固存和释放有密切联系, 其中, 溶解性有机碳是土壤微生物易利用的基质, 与有机碳矿化密切相关(刘德燕等, 2008).WSOC作为溶解性有机物组分之一, 有研究表明, WSOC与有机碳矿化速率呈极显著线性关系(李忠佩等, 2004), 而SWOC含量可用来解释土壤CO2的释放量(Marschner et al., 2002), 王艮梅等(2003)认为WSOC是微生物可直接利用的能源, 也是有机碳矿化的直接来源, 在一定条件下土壤中溶解性有机物会产生CO2(王艮梅等, 2003), Eller等(1995)也认为, 有机碳在转化为CO2、CH4之前要先转化为易被微生物分解利用的WSOC, WSOC含量增加为微生物活动提供能量的同时也增加了可利用碳源, 为微生物活动提供底物, 进而促进有机碳的矿化(胡敏杰等, 2016).本研究结果显示, 围垦养殖在一定程度上增加了沉积物WSOC含量(图 2d), 且沉积物WSOC与累积矿化量和C0呈显著正相关(p<0.01), 与以往研究结果一致(王莲阁等, 2014; 胡敏杰等, 2016), 这也进一步表明WSOC含量的高低会影响有机碳的累积矿化量, 这可能是围垦养殖塘沉积物有机碳矿化速率和累积矿化量增加的另一重要原因.另外, 沉积物有机碳累积矿化量、C0与黏粒含量呈显著正相关, 这说明沉积物粒径组成也间接影响有机碳矿化, 这主要是因为黏粒可以吸附有机质, 提高有机碳含量(李顺姬等, 2010), 进而影响有机碳矿化.
此外, 本研究结果显示, 围垦养殖总体上没有改变有机碳矿化的时间动态, 在培养前期(0~7 d), 有机碳矿化速率较快但不稳定, 变化幅度较大;培养后期(7~56 d), 矿化速率逐渐下降并趋于稳定, 这与以往室内控温实验结果相一致(李顺姬等, 2010; 胡敏杰等, 2016).出现这一结果主要与有机碳本身性质有关, 在培养前期, 沉积物中易分解碳组分被快速分解, 为微生物活动提供充足的养分, 进而有机碳的转化和CO2释放速率较快(胡敏杰等, 2016).而到了培养后期, 沉积物中易矿化分解的有机碳含量较少, 主要为难分解的有机碳, 微生物活动相对缓和(胡敏杰等, 2016), 有机碳的转化和CO2释放速率逐渐减慢, 导致有机碳矿化速率小且趋于平稳.同时, 本研究还显示, 表层沉积物有机碳矿化速率和累积矿化量均高于亚表层, 与以往多数研究结果相似(李顺姬等, 2010).不同层次有机碳矿化的差异主要由可利用性碳氮基质数量决定(李顺姬等, 2010), 本研究显示, 不同沉积物类型TOC及有机碳组分总体上表现为表层高于亚表层(图 2), 与多数研究结果相一致(张静等, 2014).张浩等(2016)认为表层沉积物接受外源有机碳的输入较多而含量提高, 而亚表层接受外来有机碳输入较少, 且深层土壤有机碳较多以惰性碳为主, 无法给微生物分解提供充足的物质来源, 所以造成亚表层有机碳矿化速率和累积矿化量低于表层, 故TOC及有机碳组分含量的差异可能是造成本研究中不同沉积层有机碳矿化速率差异的主要原因.
5 结论(Conclusions)1) 围垦养殖, 尤其是围垦15 a, 沉积物TOC和WSOC含量显著增加, MBC含量无显著变化, EOC含量则有所降低.
2) 围垦养殖促进了表层(0~10 cm)沉积物有机碳矿化速率和累积矿化量, 而亚表层(10~20 cm)沉积物有机碳矿化速率无显著差异, 且表层累积矿化量高于亚表层.
3) 养殖塘沉积物潜在矿化量(C0)高于短叶茳芏湿地, 进一步说明围垦养殖总体促进了湿地沉积物有机碳矿化, 尤其是围垦15 a养殖塘最为明显.
4) 围垦养殖塘沉积物TOC及有机碳组分的变化与长期水淹及饵料添加等管理方式有关, 同时也受黏粒、TN和TP等理化性质的间接影响, 这些变化进一步促进了沉积物有机碳矿化能力.
陈宏友, 徐国华. 2004. 江苏滩涂围垦开发对环境的影响问题[J]. 水利规划与设计, 2004(1): 18–21.
|
陈军冰, 王乘, 郑垂勇, 等. 2012. 沿海滩涂大规模围垦及保护关键技术研究概述[J]. 水利经济, 2012, 30(3): 1–5, 81.
|
Cooper J M, Burton D, Daniell T J, et al. 2011. Carbon mineralization kinetics and soil biological characteristics as influenced by manure addition in soil incubated at a range of temperatures[J]. European Journal of Soil Biology, 47(6): 392–399.
DOI:10.1016/j.ejsobi.2011.07.010
|
丁晨曦, 李永强, 董智, 等. 2013. 不同土地利用方式对黄河三角洲盐碱地土壤理化性质的影响[J]. 中国水土保持科学, 2013, 11(2): 84–89.
|
Ellert B H, Gregorich E G.1995.Management-induced Changes in the Actively Cycling Fractions of Soil Organic Matter//Mcfee W W, Kelly J M.Carbon Forms and Functions in Forest Soils[M].Wisconsin, Madison:Soil Science Society of America, Ina.119-138
|
Funge-Smith S J, Briggs M R P. 1998. Nutrient budgets in intensive shrimp ponds:implications for sustainability[J]. Aquaculture, 164(1): 117–133.
|
高灯州, 曾从盛, 章文龙, 等. 2016. 水淹频率增加对闽江口湿地土壤有机碳及其活性组分的影响[J]. 环境科学学报, 2016, 36(3): 974–980.
|
胡敏杰, 邹芳芳, 仝川, 等. 2016. 氮、硫输入对河口湿地土壤有机碳矿化的实验研究[J]. 环境科学学报, 2016, 36(11): 4184–4192.
|
Jagadamma S, Mayes M A, Steinweg J M, et al. 2014. Substrate quality alters the microbial mineralization of added substrate and soil organic carbon[J]. Biogeosciences, 11(17): 4665–4678.
DOI:10.5194/bg-11-4665-2014
|
柯跃进, 胡学玉, 易卿, 等. 2014. 水稻秸秆生物炭对耕地土壤有机碳及其CO2释放的影响[J]. 环境科学, 2014, 35(1): 93–99.
|
Laik R, Kumar K, Das D K, et al. 2009. Labile soil organic matter pools in a calciorthent after 18 years of afforestation by different plantations[J]. Applied Soil Ecology, 42(2): 71–78.
DOI:10.1016/j.apsoil.2009.02.004
|
李顺姬, 邱莉萍, 张兴昌. 2010. 黄土高原土壤有机碳矿化及其与土壤理化性质的关系[J]. 生态学报, 2010, 30(5): 1217–1226.
|
李忠佩, 张桃林, 陈碧云. 2004. 可溶性有机碳的含量动态及其与土壤有机碳矿化的关系[J]. 土壤学报, 2004, 41(4): 544–552.
DOI:10.11766/trxb200308110408 |
刘德燕, 宋长春. 2008. 磷输入对湿地土壤有机碳矿化及可溶性碳组分的影响[J]. 中国环境科学, 2008, 28(9): 769–774.
|
刘剑秋, 曾从盛, 陈宁. 2006. 闽江河口湿地研究[M]. 北京: 科学出版社: 330–334.
|
柳敏, 宇万太, 姜子绍, 等. 2007. 土壤溶解性有机碳(DOC)的影响因子及生态效应[J]. 土壤通报, 2007, 38(4): 758–764.
|
鲁如坤. 2000. 土壤化学农业分析方法[M]. 北京: 中国农业科技出版社.
|
Luo M, Zeng C S, Tong C, et al. 2014. Abundance and speciation of iron across a subtropical tidal marsh of the Min River Estuary in the East China Sea[J]. Applied Geochemistry, 45: 1–13.
DOI:10.1016/j.apgeochem.2014.02.014
|
Marschner B, Bredow A. 2002. Temperature effects on release and ecologically relevant properties of dissolves organic carbon in sterilized and biologically active soil samples[J]. Soil Biol Biochem, 34: 459–466.
DOI:10.1016/S0038-0717(01)00203-6
|
Núñez S, Martínez-Yrízar A, Búrquez A, et al. 2001. Carbon mineralization in the southern Sonoran Desert[J]. Acta Oecologica, 22(5): 269–276.
|
Ruban V, López-Sánchez J F, Pardo P, et al. 2001. Harmonized protocol and certified reference material for the determination of extractable contents of phosphorus in freshwater sediments-Asynthesis of recent works[J]. Fresenius' Journal of Analytical Chemistry, 370(2/3): 224–228.
|
余健, 房莉, 卞正富, 等. 2014. 土壤碳库构成研究进展[J]. 生态学报, 2014, 34(17): 4829–4838.
|
Tian N, Wang Y X, Weng B Q. 2010. Advances in estimating soil carbon storage[J]. Subtropical Agriculture Research, 6(3): 193–198.
|
王艮梅, 周立祥. 2003. 陆地生态系统中水溶性有机物动态及其环境学意义[J]. 应用生态学报, 2003, 14(11): 2019–2025.
DOI:10.3321/j.issn:1001-9332.2003.11.048 |
王莲阁, 高岩红, 丁长欢, 等. 2014. 变温环境对典型石灰土有机碳矿化的影响[J]. 环境科学, 2014, 35(11): 4291–4297.
|
王绍强, 于贵瑞. 2008. 生态系统碳氮磷元素的生态化学计量学特征[J]. 生态学报, 2008, 28(8): 3937–3943.
|
吴建兰. 2012. 长江入海口北支沿海滩涂养殖区底泥重金属污染特征及趋势评价[J]. 四川环境, 2012, 31(4): 76–80.
|
吴明, 邵学新, 胡锋, 等. 2008. 围垦对杭州湾南岸滨海湿地土壤养分分布的影响[J]. 土壤, 2008, 40(5): 760–764.
|
杨平, 仝川, 何清华, 等. 2013. 闽江口鱼虾混养塘水-气界面温室气体通量及主要影响因子[J]. 环境科学学报, 2013, 33(5): 1493–1503.
|
曾从盛, 钟春棋, 仝川, 等. 2008. 土地利用变化对闽江河口湿地表层土壤有机碳含量及其活性的影响[J]. 水土保持学报, 2008, 22(5): 125–129.
|
张静, 马玲, 丁新华, 等. 2014. 扎龙湿地不同生境土壤微生物生物量碳氮的季节变化[J]. 生态学报, 2014, 34(13): 3712–3719.
|
张林海, 曾从盛, 仝川. 2011. 生源要素有效性及生物因子对湿地土壤碳矿化的影响[J]. 生态学报, 2011, 31(18): 5387–5395.
|
朱丽琴, 黄荣珍, 段洪浪, 等. 2017. 红壤侵蚀地不同人工恢复林对土壤总有机碳和活性有机碳的影响[J]. 生态学报, 2017, 37(1): 249–257.
|