 2017, Vol. 37
2017, Vol. 37



2. 南京大学地球科学与工程学院, 南京 210023
2. School of Earth Sciences and Engineering, Nanjing University, Nanjing 210023
厌氧氨氧化作用是微生物以氨氮为电子供体, 将亚硝酸盐氮还原为氮气的过程(Kuypers et al., 2003).沉积物作为水环境系统中厌氧氨氧化菌富集的主要场所和氮素的重要内源, 其厌氧氨氧化脱氮过程影响水体环境氮的均衡(Dalsgaard and Thamdrup, 2002).
已有研究表明不同沉积环境介质条件下的厌氧氨氧化作用差异显著.其中, 海洋沉积环境中厌氧氨氧化脱氮贡献高达30%~50% (Devol, 2003), 河口、湿地和湖泊沉积环境厌氧氨氧化脱氮贡献分别为5%~24%(Risgaard-Petersen et al., 2004; Teixeira et al., 2014)、24%~33% (Erler et al., 2008; Zhu et al., 2011)和13%~40%(Schubert et al., 2006; Yoshinaga et al., 2011), 河流沉积环境厌氧氨氧化脱氮贡献较低, 为0.8%~10.7%(Zhao et al., 2013).近年来, 对于太湖入湖河流沉积物, Zhao等(2013)基于15N同位素配对技术开展了泥浆培养试验, 表明受水温和硝氮浓度的影响, 太湖入湖河流殷村港和河南埂沉积物的厌氧氨氧化潜在速率分别为3.08 μmol·m-2·h-1和6.79 μmol·m-2·h-1.对于太湖湖相沉积物, Wu等(2012)和Wan等(2017)基于分子生物学技术鉴别了太湖各湖区间沉积物中厌氧氨氧化菌群落存在显著差异.徐徽等(2009a)进一步结合15N同位素配对技术开展了柱状样流动培养试验, 发现太湖北部湖区梅梁湾内湾与外湾沉积物中厌氧氨氧化原位速率分别为7.50 μmol·m-2·h-1和2.05 μmol·m-2·h-1, 脱氮贡献率分别为13%和11%.西部湖区作为太湖的主要入流区, 其年均总氮入湖量约为2.68万t(胡开明等, 2012), 占太湖入湖总量的50%~65%(郑一等, 2001; 吴雅丽等, 2014).同时, 西部湖区为氮素转化活跃区(吴庆乐等, 2015), 其微生物介导下的氮转化过程极为显著.因此, 研究高氮素输入量的西部湖区沉积物厌氧氨氧化潜在速率及其脱氮贡献, 可丰富淡水系统氮的生物地球化学循环理论体系, 然而相关研究却未见报道.
本文基于15N同位素配对技术, 利用15NH4+、15NH4++14NO3-和15NO3-三组同位素开展泥浆培养试验, 评估了受外源负荷影响且高氮素输入量的太湖西部湖区沉积物四季的厌氧氨氧化潜在速率及其脱氮贡献率;并结合厌氧氨氧化功能基因及沉积物有机质、孔隙水氨氮等理化特征分析, 探讨了沉积物厌氧氨氧化潜在速率季节性差异的原因, 以期为高氮素输入湖区氮污染有效防控提供理论依据.
2 材料与方法(Materials and methods) 2.1 样品采集本研究于2013年12月和2014年6月, 在西部湖区布设了10个采样点, 采集了上覆水和沉积物样品.同时, 利用PCR-DGGE方法, 将微生物群落结构相似度大于60%的1~6号点聚为簇1, 7~10号点聚为簇2.其中, 簇1沉积物氮转化功能基因丰度约为簇2的1.3倍, 为氮转化活跃区, 并受入湖河道(南河水系)氮素输入影响.因此, 在前期研究的基础上, 于2015年9月、12月和2016年3月、6月, 在簇1中部4号点周边采集6个沉积物样品, 混合均匀作为一个均一样, 采样点位如图 1所示.采集的沉积物样品保存于可湿热灭菌气密袋中, 并置于医用酒精灭菌的4 ℃保温箱(Esky, 北京优冷)中保存, 2 h内运回实验室.分取部分沉积物, 在3000 r·min-1转速下离心30 min得到孔隙水, 经0.45 μm水系过滤膜过滤并测试其理化指标.同时, 取部分沉积物自然风干, 粉碎后过200目筛, 用于沉积物理化指标测试.
 |
| 图 1 研究区域与采集点分布示意图 (·为前期采样点) Fig. 1 Study area and sampling sites (·represents previous sampling sites) |
各季度均取45个12 mL顶空进样瓶(Labco Exetainer, Lampeter, UK), 分别加入3 g混合均匀的沉积物并加入去离子水至无顶空, 充分曝以氦气40 min达到厌氧状态, 并在水浴恒温振荡器中(200 r·min-1往复振荡)避光预培养7 d.由于湖区水深较浅且受风浪影响显著, 水体缺少温跃层, 导致湖水和浅层沉积物的温度较为接近.因此, 春、夏、秋、冬四季泥浆培养温度参照取样时湖水温度, 分别为15、25、20、10 ℃.预培养后的样品分为E_ctrl、E_amox和E_denit 3组.其中, E_ctrl作为控制组, 添加15NH4+(99% 15N), 用于判断培养体系是否稳定, 并证明耦合硝化-反硝化过程的缺失, 排除由纯化学作用反应生成N2的可能性;E_amox作为验证组, 添加15NH4+(99% 15N)+14NO3-, 用于确定厌氧氨氧化过程的存在;E_denit作为计算组, 添加15NO3-(99.19% 15N), 用于定量厌氧氨氧化潜在速率及其脱氮贡献.各试验组15N浓度均为100 μmol·L-1(表 1).在添加15N标记物后的第0、2、4、6、8 h, 每个试验组分别取3个平行样品加入150 μL的ZnCl2溶液(7 mol·L-1)终止反应, 共取得45个样品, 样品均于4 ℃低温保存以测定29N2和30N2.
| 表 1 15N泥浆培养试验组 Table 1 15N slurry incubation experiment groups |
通过测定试验组E_denit的29N2和30N2生成量, 可分别计算反硝化和厌氧氨氧化潜在速率及其脱氮贡献率(Thamdrup and Dalsgaard, 2002), 如式(1)~(4)所示.
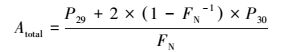
|
(1) |
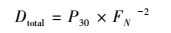
|
(2) |
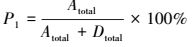
|
(3) |
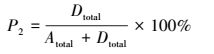
|
(4) |
式中, Atotal为厌氧氨氧化速率(μmol·kg-1·h-1);Dtotal为反硝化速率(μmol·kg-1·h-1);P1为厌氧氨氧化脱氮贡献率, P2为反硝化脱氮贡献率;FN是试验组E_denit中15NO3-占总NO3-的百分比;P29和P30分别是试验组E_denit中29N2和30N2的生成速率(μmol·kg-1·h-1).
2.3 测定方法 2.3.1 理化指标测试沉积物孔隙水中NH4+的测定参照《水和废水监测分析方法》, 以纳氏试剂光度法测定;NO3-以离子色谱仪(ICS-2000)测定.沉积物中的有机质、TN、NH4+和NO3-+NO2-的测定均参照《土壤农业化学分析方法》.其中, 有机质采用低温外热重铬酸钾氧化-比色法测定, TN以开氏消煮法测定, NH4+和NO3-+ NO2-采用MgO-代氏合金蒸馏法测定.沉积物以及孔隙水中各项理化指标的测定均采用3组平行样品.
2.3.2 功能基因鉴别分取0.5 g沉积物, 采用E.Z.N.A.TM Soil DNA Kit试剂盒(OMEGA bio-tek)提取DNA(李筱宛等, 2016).提取后的DNA样品于-20 ℃冷冻储存以进行功能基因丰度的荧光定量分析.
厌氧氨氧化功能基因(hzo)与反硝化功能基因(nirS)丰度的测定操作均在超净工作台(ZHJH-C1112B)中进行.实验采用3组平行样品, 利用SYBR Green染料法于实时荧光定量PCR仪(ABI Stepone, US)测定(Dang et al., 2010).反应体系为20 μL, 其中包含10 μL 2xSYBR Green Premix, 1.8 μL正向引物, 1.8 μL反向引物, 2 μL DNA模板(DNA样品稀释20倍), 无菌水4.4 μL (Zhang Y et al., 2016).厌氧氨氧化功能基因与反硝化功能基因所采用的引物如表 2所示(Braker et al., 1998; Hirsch et al., 2011).
| 表 2 厌氧氨氧化与反硝化功能基因引物 Table 2 Primers specific for anammox and denitrification functional genes |
培养结束后的泥浆直接于labco进样瓶中离心, 以分离泥浆为固液两相, 利用蠕动泵将上清液吸取至装有气体过滤膜系统的取样器中, 分离出溶解的气体, 最终以膜接口质谱仪(Membrane Inlet Mass Spectrometer, MIMS)直接测定可溶性气体29N2和30N2的组成和含量信息(Mccarthy and Gardner, 2003; 张波等, 2012).
3 结果与讨论(Results and discussion) 3.1 沉积环境理化特征及功能基因丰度沉积物和孔隙水理化指标如表 3所示.结果表明:沉积物有机质和孔隙水氨氮季节差异显著.西部湖区作为藻型湖区, 冬季藻类死亡沉积, 导致沉积物有机质含量显著升高.因此, 冬季沉积物有机质含量高于其他季节.然而, 西部湖区受入湖河道(南河水系)氮素的季节性输入影响, 冬季总氮输入量约为夏季的1/2(胡开明等, 2012), 导致冬季沉积物总氮含量低于夏季, 因此与有机质含量无显著的正相关性.沉积物中氮素以有机氮为主, 孔隙水中无机氮以NH4+为主(高出NO3-浓度近1个数量级), 且夏秋季含量高于春冬季, 秋季约为冬季的3倍.冬季低温抑制了沉积物有机氮的矿化与硝化作用(周才平等, 2001), 减少了孔隙水中无机氮的内源释放量, 导致孔隙水NH4+和NO3-均明显低于其他季节.
| 表 3 沉积物、孔隙水和上覆水理化指标 Table 3 The physico-chemical characters of sediments, pore water and overlying water |
沉积物厌氧氨氧化(hzo)和反硝化(nirS)功能基因丰度季节差异显著, 如表 4所示.沉积物反硝化功能基因丰度明显高出厌氧氨氧化功能基因1~2个数量级.同时, 夏秋季沉积物的厌氧氨氧化功能基因丰度高于春冬季, 秋季最高, 为9.70×107copies·g-1, 冬季最低, 为2.30×106 copies·g-1.
| 表 4 沉积物nirS和hzo丰度值 Table 4 Abundance of nirS and hzo in sediments |
3组泥浆培养试验产生的15N结果如图 2所示.其中, E_ctrl组中各季度的样品均未明显生成29N2和30N2, 表明泥浆中NO3-背景值降低为零, 缺失耦合硝化-反硝化过程, 培养体系达到稳定状态, 而且排除了由纯化学作用(如MnO2氧化NH4+)生成N2的可能(Luther et al., 1997);试验组E_amox中各季度的泥浆产生的29N2均存在显著的增长趋势, 表明泥浆中存在厌氧氨氧化过程;试验组E_denit中各季度泥浆产生的30N2和29N2都显著增加, 且30N2产生速率明显快于29N2, 表明四季沉积物均存在反硝化与厌氧氨氧化过程, 且反硝化速率大于厌氧氨氧化潜在速率.
 |
| 图 2 泥浆培养试验 29N2、30N2生成量 (图中E_ctrl-xN2、E_amox-xN2、E_denit-xN2分别表示E_ctrl、E_amox、E_denit试验组的29N2、30N2生成量) Fig. 2 The yield of 29N2 and 30N2slurry incubation experiment groups (E_ctrl-xN2, E_amox-xN2 and E_denit-xN2 are the yield of 29N2 and 30N2 of E_ctrl, E_amox and E_denit groups respectively) |
试验组E_denit中, 29N2和30N2生成量随时间线性增加(图 2), 根据其斜率可得到29N2和30N2的产生速率, 分别以P29和P30表示.根据P29、P30, 由公式(1)、(2)可分别计算厌氧氨氧化和反硝化潜在速率Atotal和Dtotal, 进一步由公式(3)、(4)得到相应的脱氮贡献率P1和P2, 计算结果见表 5.研究区域四季沉积物厌氧氨氧化潜在速率为3.24~10.11 μmol·kg-1·h-1, 最大值和最小值分别出现在秋季和冬季, 其季节变化差异可达3~4倍;脱氮贡献率的季节差异性不显著, 为30%~40%, 远高于梅梁湾沉积物(11%~14%)(徐徽等, 2009a).其主要原因为西部湖区厌氧氨氧化功能基因丰度为2.30×106~9.70×107 copies·g-1, 显著高于梅梁湾的厌氧氨氧化功能基因丰度(6.82×106~1.33×107 copies·g-1, 未发表).同时, 西部湖区沉积物氨氮释放速率高于梅梁湾(徐徽等, 2009b), 提供厌氧氨氧化反应的底物越多, 厌氧氨氧化脱氮效应越高.
| 表 5 四季反硝化、厌氧氨氧化潜在速率及其贡献 Table 5 Rates and the contribution to the nitrogen loss of anammox and denitrification |
已有研究表明, 温度可通过影响膜转运蛋白的亲和性而改变微生物代谢能力(Khalil and Richards, 2011; Qiang et al., 2016), 低温可降低微生物对基质的可利用性, 导致厌氧氨氧化功能基因丰度值降低, Li等(2011)发现当温度从30 ℃降低至20 ℃时, 厌氧氨氧化功能基因丰度可降低近1个数量级.氨氮作为厌氧氨氧化过程的电子供体和微生物生长基质, 其浓度的增加会导致厌氧氨氧化功能基因丰度的增加(Hu et al., 2012).本研究中, 厌氧氨氧化功能基因丰度夏秋季明显高于春冬季, 且与孔隙水氨氮表现出相似的季节性变化特征(图 3a), 表明温度、孔隙水氨氮是影响厌氧氨氧化功能基因丰度的主要原因.此外, 溶解氧及氧化还原电位(ORP)决定着氧化还原的反应方向, 有研究表明沉积物厌氧氨氧化功能基因丰度随上覆水溶解氧浓度增加而降低(付璐璐等, 2016).研究区域夏秋季藻类生长迅速, 造成水体中溶解氧浓度下降(陈晓峰等, 2011), 导致上覆水溶解氧以及氧化还原电位(ORP)明显低于春冬季(表 3), 进而引起沉积物中厌氧氨氧化功能基因丰度的增加.同时, pH值对厌氧氨氧化菌存在显著影响, 研究表明厌氧氨氧化菌的最适宜pH值为8左右(杨洋等, 2006), 西部湖区上覆水pH值四季差异不显著且均接近8, 因此本研究中上覆水pH值不是厌氧氨氧化功能基因丰度季节变化的主要影响因素.
 |
| 图 3 孔隙水NH4+(a)和厌氧氨氧化速率Atotal(b)和hzo基因丰度值季节分布 Fig. 3 The seasonal variation of NH4+ concentration in pore water(a) and Atotal(b) and abundance of hzo in sediment |
同时, 厌氧氨氧化功能基因丰度的增加会提高厌氧氨氧化潜在速率(Hou et al., 2013), 因此环境因子可通过改变厌氧氨氧化功能基因丰度(Rich et al., 2008; Wang et al., 2012), 进而影响厌氧氨氧化脱氮能力(Guo et al., 2015).本研究显示夏秋季沉积物中厌氧氨氧化功能基因丰度和厌氧氨氧化潜在速率均显著高于春冬季(图 3b), 表明夏秋季较高温度和孔隙水氨氮浓度、较低上覆水溶解氧浓度和氧化还原电位可通过提高厌氧氨氧化功能基因丰度, 进而促进厌氧氨氧化潜在速率.
此外, 泥浆样品本底亚硝氮含量低于检测限(<0.03 mg·L-1), 厌氧氨氧化过程所需的电子受体(亚硝氮)来源于反硝化过程的第一步.因此, 反硝化与厌氧氨氧化对亚硝氮的利用存在竞争, 从而影响厌氧氨氧化潜在速率及其脱氮贡献(徐徽等, 2009a).同时, 也有研究表明较高的有机质含量则会直接抑制厌氧氨氧化菌, 导致厌氧氨氧化潜在速率明显下降(Güven et al., 2005; 张诗颖等, 2015).其中, 反应器中高有机质的载入(95~400 mg·COD·L-1)会逐步抑制厌氧氨氧化菌活性, 当COD超过300 mg·L-1后, 厌氧氨氧化菌会表现为无活性甚至死亡(Waki et al., 2007; Chamchoi et al., 2008; Molinuevo et al., 2009).研究区域沉积物有机质含量春冬季高于夏秋季(表 3), 会促进以有机碳为电子供体的反硝化脱氮过程, 增加反硝化对亚硝氮的利用率, 间接抑制厌氧氨氧化脱氮过程(图 4).因此, 研究区域春冬季沉积物高含量有机质是抑制厌氧氨氧化潜在速率的重要原因.
 |
| 图 4 有机质含量与厌氧氨氧化速率Atotal季节变化 Fig. 4 The seasonal variation of OM content and Atotal in the sediment |
1) 沉积物有机质含量、孔隙水氨氮浓度和厌氧氨氧化功能基因丰度季节差异显著.其中, 有机质含量为11.29~39.32 mg·g-1, 夏秋季低于春冬季;孔隙水氨氮浓度为0.73~3.16 mg·L-1, 厌氧氨氧化功能基因丰度为2.30×106~9.70×107 copies·g-1, 两者均表现为夏秋季高于春冬季.
2) 厌氧氨氧化潜在速率季节差异性显著, 夏秋季显著高于春冬季, 差异可达3~4倍.其中, 厌氧氨氧化潜在速率最大值和最小值分别出现在秋季和冬季, 其值分别为10.11 μmol·kg-1·h-1和3.24 μmol·kg-1·h-1.同时, 沉积物四季总脱氮速率为10.67~31.02 μmol·kg-1·h-1, 厌氧氨氧化作用脱氮效应显著, 四季脱氮贡献率分别为36%、30%、40%和30%.
3) 较高温度和孔隙水氨氮浓度、较低上覆水溶解氧浓度和氧化还原电位导致厌氧氨氧化功能基因丰度升高, 进而促进厌氧氨氧化脱氮过程.有机质作为反硝化过程的电子供体, 会增加反硝化对亚硝氮的利用率, 进而抑制厌氧氨氧化脱氮过程.因此, 推断夏秋季较高厌氧氨氧化功能基因丰度和较低的沉积物有机质含量是导致其厌氧氨氧化潜在速率显著高于春冬季的重要原因.
Braker G, Fesefeldt A, Witzel K P, et al. 1998. Development of PCR primer systems for amplification of nitrite reductase genes (nirK and nirS) to detect denitrifying bacteria in environmental samples[J]. Applied and Environmental Microbiology, 64(10): 3769–3775.
|
Chamchoi N, Nitisoravut S, Schmidt J E. 2008. Inactivation of anammox communities under concurrent operation of anaerobic ammonium oxidation (anammox) and denitrification[J]. Bioresource Technology, 99(9): 3331–3336.
DOI:10.1016/j.biortech.2007.08.029
|
陈晓峰, 周伟杰, 钮伟民, 等. 2011. 环太湖流域藻类密度和主要污染指标的相关性研究[J]. 环境与健康杂志, 2011, 28(5): 422–424.
|
Dalsgaard T, Thamdrup B. 2002. Factors controlling anaerobic ammonium oxidation with nitrite in marine sediments[J]. Applied and Environmental Microbiology, 68(8): 3802–3808.
DOI:10.1128/AEM.68.8.3802-3808.2002
|
Dang H, Li J, Chen R, et al. 2010. Diversity, abundance, and spatial distribution of sediment ammonia-oxidizing betaproteobacteria in response to environmental gradients and coastal eutrophication in Jiaozhou Bay, China[J]. Applied and Environmental Microbiology, 76(14): 4691–4702.
DOI:10.1128/AEM.02563-09
|
Devol A H. 2003. Nitrogen cycle: Solution to a marine mystery[J]. Nature, 422(6932): 575–576.
DOI:10.1038/422575a
|
Erler D V, Eyre B D, Davison L. 2008. The contribution of anammox and denitrification to sediment N2 production in a surface flow constructed wetland[J]. Environmental Science and Technology, 42(24): 9144–9150.
DOI:10.1021/es801175t
|
付璐璐, 甄毓, 贺惠, 等. 2016. 长江口邻近海域沉积物中厌氧氨氧化细菌分布特征研究[J]. 环境科学, 2016, 37(10): 3914–3922.
|
Guo Q, Xing B S, Li P, et al. 2015. Anaerobic ammonium oxidation (anammox) under realistic seasonal temperature variations: Characteristics of biogranules and process performance[J]. Bioresource Technology, 192: 765–773.
DOI:10.1016/j.biortech.2015.06.049
|
Güven D, Dapena A, Kartal B, et al. 2005. Propionate oxidation by and methanol inhibition of anaerobic ammonium-oxidizing bacteria[J]. Applied and Environmental Microbiology, 71(2): 1066–1071.
DOI:10.1128/AEM.71.2.1066-1071.2005
|
Hirsch M D, Long Z T, Song B. 2011. Anammox bacterial diversity in various aquatic ecosystems based on the detection of hydrazine oxidase genes (hzoA/hzoB)[J]. Microbial Ecology, 61(2): 264–276.
DOI:10.1007/s00248-010-9743-1
|
Hou L, Zheng Y, Liu M, et al. 2013. Anaerobic ammonium oxidation (anammox) bacterial diversity, abundance, and activity in marsh sediments of the Yangtze Estuary[J]. Journal of Geophysical Research, 118(3): 1237–1246.
|
Hu B L, Shen L D, Zheng P, et al. 2012. Distribution and diversity of anaerobic ammonium-oxidizing bacteria in the sediments of the Qiantang River[J]. Environmental Microbiology Reports, 4(5): 540–547.
DOI:10.1111/emi4.2012.4.issue-5
|
胡开明, 逄勇, 王华. 2012. 太湖湖体总氮平衡及水质可控目标[J]. 水科学进展, 2012, 23(4): 555–562.
|
Khalil M I, Richards K G. 2011. Denitrification enzyme activity and potential of subsoils under grazed grasslands assayed by membrane inlet mass spectrometer[J]. Soil Biology and Biochemistry, 43(9): 1787–1797.
DOI:10.1016/j.soilbio.2010.08.024
|
Kuypers M M, Sliekers A O, Lavik G, et al. 2003. Anaerobic ammonium oxidation by anammox bacteria in the Black Sea[J]. Nature, 422(6932): 608–611.
DOI:10.1038/nature01472
|
Li M, Cao H, Hong Y G, et al. 2011. Seasonal dynamics of anammox bacteria in estuarial sediment of the Mai Po Nature Reserve revealed by analyzing the 16S rRNA and hydrazine oxidoreductase (hzo) genes[J]. Microbes and Environments, 26(1): 15–22.
DOI:10.1264/jsme2.ME10131
|
李筱宛, 张亚平, 万宇, 等. 2016. 上覆水不同C/N比条件下沉积物环境反硝化及氨氧化功能基因丰度变化特征[J]. 南京大学学报(自然科学), 2016, 52(1): 27–35.
|
Luther G W, Sundby B, Lewis B L, et al. 1997. Interactions of manganese with the nitrogen cycle: Alternative pathways to dinitrogen[J]. Geochimica Et Cosmochimica Acta, 61(19): 4043–4052.
DOI:10.1016/S0016-7037(97)00239-1
|
Mccarthy M J, Gardner W S. 2003. An application of membrane inlet mass spectrometry to measure denitrification in a recirculating mariculture system[J]. Aquaculture, 218(1/4): 341–355.
|
Molinuevo B, Garcia M, Karakashev D, et al. 2009. Anammox for ammonia removal from pig manure effluents: effect of organic matter content on process performance[J]. Bioresource Technology, 100(7): 2171–2175.
DOI:10.1016/j.biortech.2008.10.038
|
Qiang L, Vrieze J D, Li J, et al. 2016. Temperature affects microbial abundance, activity and interactions in anaerobic digestion[J]. Bioresource Technology, 209: 228–236.
DOI:10.1016/j.biortech.2016.02.132
|
Rich J J, Dale O R, Song B, et al. 2008. Anaerobic ammonium oxidation (anammox) in Chesapeake Bay sediments[J]. Microbial Ecology, 55(2): 311–320.
DOI:10.1007/s00248-007-9277-3
|
Risgaard-Petersen N, Meyer R L, Schmid M, et al. 2004. Anaerobic ammonium oxidation in an estuarine sediment[J]. Aquatic Microbial Ecology, 36(3): 293–304.
|
Schubert C J, Durisch-Kaiser E, Wehrli B, et al. 2006. Anaerobic ammonium oxidation in a tropical freshwater system (Lake Tanganyika)[J]. Environmental Microbiology, 8(10): 1857–1863.
DOI:10.1111/emi.2006.8.issue-10
|
Teixeira C, Magalhães C, Joye S B, et al. 2014. The contribution of anaerobic ammonium oxidation to nitrogen loss in two temperate eutrophic estuaries[J]. Estuarine Coastal and Shelf Science, 143(4): 41–47.
|
Thamdrup B, Dalsgaard T. 2002. Production of N2 through anaerobic ammonium oxidation coupled to nitrate reduction in marine sediments[J]. Applied and Environmental Microbiology, 68(3): 1312–1318.
DOI:10.1128/AEM.68.3.1312-1318.2002
|
Waki M, Tokutomi T, Yokoyama H, et al. 2007. Nitrogen removal from animal waste treatment water by anammox enrichment[J]. Bioresource Technology, 98(14): 2775–2780.
DOI:10.1016/j.biortech.2006.09.031
|
Wan Y, Ruan X H, Zhang Y P, et al. 2017. Illumina sequencing-based analysis of sediment bacteria community in different trophic status freshwater lakes[J]. Microbiology Open, 6(4): 1–15.
|
Wang S, Zhu G, Peng Y, et al. 2012. Anammox bacterial abundance, activity, and contribution in riparian sediments of the Pearl River estuary[J]. Environmental Science and Technology, 46(16): 8834–8842.
DOI:10.1021/es3017446
|
吴庆乐, 阮晓红, 吴朝明, 等. 2015. 太湖西部河湖氮污染物来源及转化途径分析[J]. 环境科学学报, 2015, 35(12): 3883–3889.
|
吴雅丽, 许海, 杨桂军, 等. 2014. 太湖水体氮素污染状况研究进展[J]. 湖泊科学, 2014, 26(1): 19–28.
DOI:10.18307/2014.0103 |
Wu Y C, Xiang Y, Wang J J, et al. 2012. Molecular detection of novel anammoxbacterial clusters in the sediments of the shallow freshwater Lake Taihu[J]. Geomicrobiology Journal, 29(9): 852–859.
DOI:10.1080/01490451.2011.635760
|
徐徽, 张路, 商景阁, 等. 2009a. 太湖梅梁湾水土界面反硝化和厌氧氨氧化[J]. 湖泊科学, 2009a, 21(6): 775–781.
|
徐徽, 张路, 商景阁, 等. 2009b. 太湖水土界面氮磷释放通量的流动培养研究[J]. 生态与农村环境学报, 2009b, 25(4): 66–71.
|
杨洋, 左剑恶, 沈平, 等. 2006. 温度、pH值和有机物对厌氧氨氧化污泥活性的影响[J]. 环境科学, 2006, 27(4): 691–695.
|
Yoshinaga I, Amano T, Yamagishi T, et al. 2011. Distribution and diversity of anaerobic ammonium oxidation (anammox) bacteria in the sediment of a eutrophic freshwater lake, Lake Kitaura, Japan[J]. Microbes and Environments, 26(3): 189–197.
DOI:10.1264/jsme2.ME10184
|
张波, 杜应旸, 陈宇炜, 等. 2012. 太湖流域典型河流沉积物的反硝化作用[J]. 环境科学学报, 2012, 32(8): 1866–1873.
|
张诗颖, 吴鹏, 宋吟玲, 等. 2015. 厌氧氨氧化与反硝化协同脱氮处理城市污水[J]. 环境科学, 2015, 36(11): 4174–4179.
|
Zhang Y, Ruan X, Wan Y, et al. 2016. Effects of Environmental Factors on Anammox Bacterial Community Structure in Sediments of a Freshwater Aquaculture Farm, Yangcheng Lake[J]. Geomicrobiology Journal, 33(6): 479–487.
DOI:10.1080/01490451.2015.1054005
|
Zhao Y, Xia Y, Kana T M, et al. 2013. Seasonal variation and controlling factors of anaerobic ammonium oxidation in freshwater river sediments in the Taihu Lake region of China[J]. Chemosphere, 93(9): 2124–2131.
DOI:10.1016/j.chemosphere.2013.07.063
|
郑一, 王学军, 江耀慈, 等. 2001. 环太湖河道水质分析与入湖污染物负荷量估算[J]. 地理学与国土研究, 2001, 17(1): 40–44.
|
周才平, 欧阳华. 2001. 长白山两种主要林型下土壤氮矿化速率与温度的关系[J]. 生态学报, 2001, 21(9): 1469–1473.
|
Zhu G, Wang S, Feng X, et al. 2011. Anammox bacterial abundance, biodiversity and activity in a constructed wetland[J]. Environmental Science and Technology, 45(23): 9951–9958.
DOI:10.1021/es202183w
|