 2017, Vol. 37
2017, Vol. 37



镍(Ni)不仅是作物生长必需的微量元素,同时也是对作物有毒的元素(刘国栋,2001).适量Ni有利于作物生长(朱京涛等,2008;杨红超等,2011),但过量Ni则对作物有毒害作用(李雪等,2015;Molas,1998),同时还可通过食物链危及人类健康(王夔,1992).有关土壤外源Ni对小麦作物的生物毒性和农田土壤阈值的研究国内已有相关报道,例如,白英等(2014)研究发现,土壤中添加Ni临界值为240 mg·kg-1时,各器官干重平均减少11.60%;张素霞等(1991)研究发现,盆栽实验小麦受害的Ni投加量为150 mg·kg-1;蒋德福等(1987)的研究表明,盆栽小麦土壤中添加Ni量高于200 mg·kg-1时会对小麦产生明显的抑制作用;黄锦孙等(2012)研究提出,祁阳土壤外源Ni对小麦籽粒的EC10为41.00 mg·kg-1,当祁阳土壤外源Ni高于100 mg·kg-1时小麦全部死亡.试验结果表明,造成小麦作物Ni毒性临界值差异的原因与供试土壤性质和作物品种不同所表现的忍耐力和抗性差异有关.青长乐等(1992)研究提出,各种作物对重金属的反应,不同生育期对重金属的敏感程度和中毒反应有所不同,并提出以作物的苗期最为敏感.笔者已对利用小麦三叶期毒害效应指标表征Ni毒害临界值进行了探讨(任艳军等,2017),杨林书等(1996)运用小麦幼苗(三叶期)毒性指标,提出并表征了土壤Cd污染的毒性临界值.已有的研究结果表明(任艳军等,2016),小麦三叶期到孕穗期Ni主要积累于根系,进入成熟期后Ni主要积累于茎秆和籽粒中,拔节期至孕穗期是Ni元素吸收速率最高的关键时期.由此看出,土壤外源Ni污染环境下,小麦孕穗期Ni的丰缺直接影响到植株的生长发育和籽实产量,实现小麦作物中期(孕穗期)重金属毒害诊断具有重要意义.而目前研究小麦孕穗期Ni毒害效应与成熟期籽实产量之间关系的系统研究尚未见报道.因此,本试验以春小麦品种“冀张春3号”为试材,采用盆栽试验方法,研究春小麦孕穗期毒害效应与成熟期籽实产量的变化及其相关关系,实现外源Ni胁迫条件下小麦作物资源毒害效应的中期诊断和评价,为小麦作物栽培和土壤资源利用提供科学方法和依据.
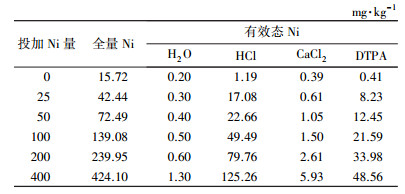
2 材料和方法(Materials and methods) 2.1 供试土壤和指示作物背景土壤采自河北科技师范学院作物试验站,土壤基本理化特性见表 1.试验土壤采用人工模拟方法,外源Ni投加量分别为0、25、50、100、200、400 mg·g-1,添加Ni盐为NiSO4·6H2O,同时按N 0.2 mg·g-1、P2O5 0.1 mg·g-1和K2O 0.1 mg·g-1的土施肥量将氮、磷、钾肥施入试验土壤中作为底肥,施入肥料分别为CO(NH2)2、KH2PO4和K2SO4,将Ni盐和化学肥料溶解后向背景土壤中投加,混合均匀后厌氧培养30 d至吸附达到平衡.然后测定土壤中全量Ni和有效态Ni(4种提取试剂)的含量,测定结果见表 2.供试作物品种为春小麦品种—冀张春3号.
| 表 1 背景土壤基本理化特性 Table 1 Physical and chemical properties of background soil |

| 表 2 外源Ni、土壤全量Ni和有效态Ni含量 Table 2 Exogenous Ni, total Ni and available Ni contents in soilmg·kg-1 |
试验在具有遮雨设施的网室内自然光照条件下进行,将充分混合后的各处理剂量土壤分别装入28 cm×40 cm的营养瓷盆中,每盆为一小区,每剂量处理3次重复.于2014年3月13日直播春小麦种子,每盆播种50~60粒,待第一片真叶长出后定苗,每盆35株.按常规要求管理,土壤湿度保持田间持水量的75%±2%.分别于小麦三叶期、拔节期、孕穗期、乳熟期、成熟期采集小麦全植株样品,每盆3株,并将植株的根系、茎杆、叶片、穗(颖壳)、籽粒等组织器官分解,分别测定其鲜质量,经杀青(籽粒80 ℃,根、茎、叶105 ℃)处理后,然后在65 ℃或70 ℃恒温条件下烘干至恒重,并称其干质量.样品经研磨处理后,存放于自封袋中待测.
2.3 分析方法土壤和植株全Ni含量测定分别采用王水-HClO4(V(HCl):V(HNO3):V(HClO4)=3:1:1) 和HNO3-HClO4(V(HNO3):V(HClO4)=4:1) 混合酸消化处理;土壤有效态Ni含量测定采用马建军等(2006)研究分析方法进行,即分别采用H2O、HCl、CaCl2、DTPA进行浸提处理.提取液:H2O(去离子水),0.1 mol·L-1 HCl,0.1 mol·L-1 CaCl2(pH=7.0),0.005 mol·L-1 DTPA-0.1 mol·L-1 TEA-0.01 mol·L-1 CaCl2溶液(pH=7.30);提取时间:DTPA为2 h,H2O、HCl和CaCl2均为60 min;土液比:m(土壤):V(DTPA)=1 g:2 mL,m(土壤):V(H2O、HCl、CaCl2)=1 g:5 mL,提取温度:均为恒温(25±1) ℃,往复振荡频率为180次·min-1.土壤和植株样品处理液分析均采用Optima2100DV等离子体发射光谱仪.
Ni生物富集系数:生物富集系数是衡量土壤重金属被植物吸收难易程度的指标,其值越大表示该重金属越易被植物吸收,计算公式见式(1).
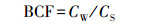
|
(1) |
式中,Cw为小麦某一器官中Ni含量(mg·kg-1),CS为土壤中全Ni量(mg·kg-1).
2.4 数据处理数据方差分析采用DPS7.05 Duncan新复极差法进行差异性显著分析,采用SPSS17.00软件进行数据相关分析.
3 结果(Results) 3.1 外源Ni对小麦孕穗期生长发育和成熟期籽实产量的影响外源Ni会增加小麦孕穗期地上部植株高度和叶片干质量(表 3).随施Ni量增加均呈先增后降趋势,50 mg·kg-1Ni处理株高增至最大,较对照增加13.84%(p < 0.05),100 mg·kg-1Ni处理叶片干质量增至最大,较对照增加7.81%(p>0.05),之后随施Ni量增加呈递减趋势;外源Ni会降低小麦植株茎秆和麦穗干质量,25和50 mg·kg-1Ni处理茎秆干质量较对照下降不明显,100、200和400 mg·kg-1Ni处理呈明显下降,较对照分别降低了18.58%、20.06%和28.91%(p < 0.05).而麦穗干质量在施Ni量25~100 mg·kg-1范围内未受明显影响,200和400 mg·kg-1Ni处理抑制程度较大,较对照分别降低了17.24%和33.62%(p < 0.05).试验表明,外源Ni对小麦孕穗期各组织器官生理代谢的影响存在一定差异.
| 表 3 外源Ni对小麦孕穗期生长发育和成熟期籽实产量的影响 Table 3 Effects of exogenous Ni on wheat development at booting stage and yield at maturity stage |
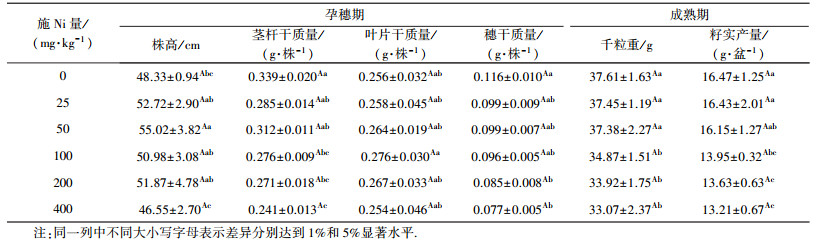
由于外源Ni对小麦孕穗期各组织器官的生长发育产生影响,势必影响小麦成熟期生殖器官籽实的营养物质代谢和干物质积累(表 3).外源Ni抑制了小麦籽实干物质的积累,使小麦籽实千粒重和单区产量指标降低,且表现为低剂量产量略下降,高剂量产量下降明显.其中,25、50 mg·kg-1Ni处理籽实千粒重和单区产量较对照分别下降0.43%、0.61%(p>0.05) 和0.22%、1.94%(p>0.05);随投加Ni量的增加,100、200、400 mg·kg-1Ni处理籽实千粒重和单区产量较对照分别下降7.29%、9.81%、12.07%(p < 0.05) 和15.32%、17.26%、19.77%(p < 0.05).各处理籽实产量(千粒重+单区产量)降幅均值分别为0.32%、1.28%、11.30%、13.54%和15.92%.表明外源Ni对小麦籽实产量的影响与孕穗期相关组织器官生长发育Ni毒害抑制程度有关.
3.2 外源Ni对小麦孕穗期各器官和成熟期籽实中Ni吸收富集的影响由图 1a可见,背景土壤条件下,小麦孕穗期植株各器官中Ni含量由大到小顺序为:根系(19.16 mg·kg-1)>叶片(3.17 mg·kg-1) >茎秆(2.90 mg·kg-1) >穗(1.58 mg·kg-1).外源Ni明显增加土壤中全量Ni和有效态Ni含量(表 2),使小麦孕穗期各器官中Ni含量呈明显增加,除根系中Ni含量呈持续明显增加外(p < 0.01),其它各器官Ni吸收出现二次吸收高峰,25 mg·kg-1处理以下出现一次吸收高峰,25~50 mg·kg-1处理,各器官中Ni含量由大到小顺序为:根系(86.69~149.31 mg·kg-1)>穗(6.34~6.53 mg·kg-1)>茎秆(6.00~6.12 mg·kg-1)>叶片(4.58~4.56 mg·kg-1),各器官中Ni含量明显高于对照处理(p < 0.01);超过100 mg·kg-1处理出现二次吸收高峰,各器官中Ni含量呈明显增加(p < 0.01),100~400 mg·kg-1处理,Ni含量由大到小顺序为:根系(318.49~1128.02 mg·kg-1)>穗(19.17~63.67 mg·kg-1)>茎秆(10.41~54.88 mg·kg-1)>叶片(7.35~38.70 mg·kg-1).由此可见,外源Ni会改变小麦孕穗期各器官中Ni的吸收与积累,除根系中Ni含量明显高于其它各器官外,穗中Ni的含量高于营养器官茎秆和叶片.造成小麦孕穗期各器官Ni吸收差异与Ni富集能力大小变化密切相关,图 1b结果显示,背景土壤条件下,各器官Ni富集系数大小与Ni含量分布一致,与背景土壤比较,外源Ni增加了根系和穗中Ni的富集系数(除50 mg·kg-1处理外),降低了茎秆和叶片中Ni的富集系数;且随投加Ni量增加,根系中Ni富集系数呈持续递增,穗中Ni富集系数出现二次高峰,而茎秆和叶片中Ni富集系数呈明显下降,超过200 mg·kg-1处理Ni富集系数回升,Ni富集能力由大到小的顺序为:根系>穗>茎秆>叶片,其中,根系Ni富集系数大于1,其它各器官Ni富集系数均明显小于1.表明小麦孕穗期根系吸收富集Ni能力最强,与文献报道(任艳军等,2016)的小麦孕穗期Ni吸收速率最快,根系Ni累积量最高(分配率占47.93%)的结果相吻合.由此可见,外源Ni改变了小麦孕穗期各组织器官中Ni的吸收与再分配,Ni含量的变化可能是影响各器官干物质积累的重要因子.
 |
| 图 1 外源Ni对小麦孕穗期各器官中Ni含量的影响(a)及Ni富集系数的影响(b) Fig. 1 Effects of exogenous Ni on Ni contents (a) and Ni bioaccumulation (b) in wheat organs at booting stage |
外源Ni对小麦成熟期籽实中Ni含量及Ni富集系数的影响与孕穗期穗中相应指标变化规律一致,也同样出现二次吸收和富集高峰(图 2),表明小麦孕穗期至成熟期生殖器官营养物质代谢具有遗传稳定性.其中,背景土壤条件下,小麦籽实Ni含量为1.43 mg·kg-1,25和50 mg·kg-1处理,籽实Ni含量分别为7.39和8.05 mg·kg-1,两处理之间Ni含量无明显差异(p>0.05),100、200和400 mg·kg-1处理,籽实Ni含量分别为18.00、38.47和79.82 mg·kg-1 (p < 0.01).试验结果表明,小麦生殖器官具有较高的Ni富集能力,是造成生殖器官中Ni含量高于营养器官中的重要因子.鉴于此,对于小麦作物食用器官中Ni含量的分布规律研究应予以重视.
 |
| 图 2 外源Ni对小麦成熟期籽粒中Ni含量(a)及Ni富集系数(b)的影响 Fig. 2 Effects of exogenous Ni on Ni contents (a) and Ni bioaccumulation (b) in wheat grains at maturity stage |
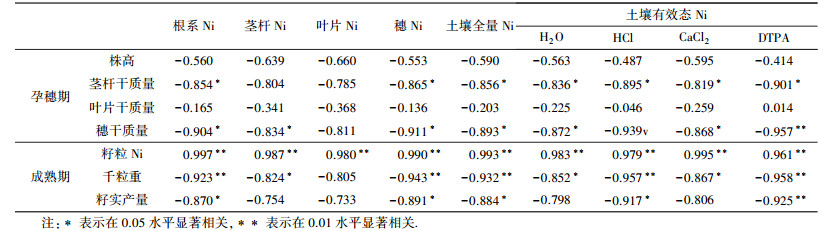
试验结果显示(表 2),外源Ni会增加土壤中不同形态Ni的含量,造成小麦孕穗期各器官和成熟期籽实中Ni的吸收积累,二者之间相关分析结果表现为呈显著正相关关系(r=0.957~0.998,p < 0.01);而小麦孕穗期根系、穗中Ni含量和土壤中不同形态Ni含量分别与茎秆干质量、穗干质量均表现显著负相关(p < 0.05),表明土壤不同形态Ni及根系、穗中Ni在影响地上部茎秆和穗中干物质积累方面关系密切,造成小麦孕穗期各器官干物质质量变化差异可能与相应器官中Ni毒害作用大小有关.
为探求小麦孕穗期各器官生长发育与成熟期籽实产量Ni毒害效应的关系,实现小麦籽实产量Ni毒害效应的中期诊断,进行小麦孕穗期和成熟期生理指标间相关分析.结果显示(表 4),小麦根系和穗中Ni含量及土壤中全量Ni和有效态Ni(HCl和DTPA提取态)与小麦籽实千粒重和籽实产量均达到显著负相关(p < 0.05, p < 0.01).同时,小麦孕穗期茎秆和穗干质量均与成熟期籽实产量和千粒重呈显著正相关关系(表 5).试验结果表明,小麦孕穗期和成熟期外源Ni毒害效应具有持续性和一致性,利用小麦孕穗期相关器官生理指标变化可以实现外源Ni对小麦成熟期产量指标毒害效应的中期诊断与评价.
| 表 4 外源Ni与小麦孕穗期和成熟期各器官生理指标间的相关系数 Table 4 Correlation analysis between exogenous Ni and wheat physiological indexes at different stages |
| 表 5 小麦孕穗期生理指标与成熟期籽实产量间的相关系数 Table 5 Correlation analysis between wheat physiological indexes at booting stage and yield at maturity stage |
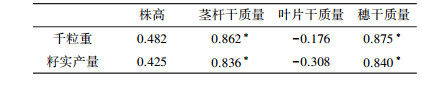
Ni是植物必需的营养元素,是植物脲酶、一氧化碳脱氢酶和甲醛还原酶等酶蛋白的金属辅基,是植物氮代谢必不可少的营养元素(Eskew et al., 1983).适量Ni可促进植物氮代谢,有利于植物干物质的积累和生长发育(Patrick et al., 1987);过量Ni会降低叶绿素含量,使光合作用能力减弱,不利于光合代谢产物的积累与贮存,进而产生毒害抑制作用(Gajewska et al., 2006).本研究表明,该试验浓度条件下,外源Ni增加了小麦孕穗期植株株高,促进了叶片部位的干物质的积累,对小麦茎秆和穗部位产生了抑制作用,使干物质积累下降(表 3),表明外源Ni对小麦不同部位产生的毒害效应存在差异.研究报道,土壤-植物系统中重金属的积累能力和生物毒性不但与其总量有关,在很大程度上还由其形态分布决定,不同形态产生的环境效果不同(Ramos et al., 1994).一般认为,可交换态和水溶态重金属是植物可吸收利用的主要形态(Wang et al., 1997),可交换态Ni更易于被小麦吸收和转移(白英等,2014).本研究表明,外源Ni会增加土壤中有效态Ni含量,随Ni投加量的增加,小麦各器官中Ni含量随之递增,土壤Ni含量与小麦各器官中Ni含量之间均表现显著的正相关关系(r=0.957~0.998,p < 0.01).本文结果显示,小麦孕穗期地上部各器官不同剂量处理Ni含量均值由大到小顺序为:穗(26.94 mg·kg-1)>茎秆(19.35 mg·kg-1)>叶片(13.61 mg·kg-1);Ni含量增幅均值由大到小顺序为:穗(1605.29%)>茎秆(567.23%)>叶片(329.24%);Ni富集系数均值由大到小顺序为:穗(0.14)>茎秆(0.10)>叶片(0.07).由此可见,由于各器官吸收累积Ni量和富集能力的差异,造成各器官Ni毒害效应的差异,小麦各器官干质量减产幅度均值由大到小顺序为:穗(-21.38%)>茎秆(-18.29%)>叶片(3.05%).研究发现,小麦地上部各器官的毒害效应程度高低与Ni含量高低关系密切,Ni含量越高,毒害作用能力越强,表明小麦植株外源Ni毒害效应主要是由Ni元素积累富集毒害所致.
研究发现,Ni能在植物有生长力的部位与再生殖部位之间迁移和积累(Seregin et al., 2007).试验结果表明,同样Ni添加量下,小麦孕穗期地上部生殖器官中Ni富集系数明显高于其它营养器官,穗中Ni含量和增幅明显高于茎秆和叶片部位,小麦生殖器官对Ni吸收富集能力强于其它各器官,表明小麦进入生殖阶段(孕穗期至成熟期)Ni更易向生殖器官中富集积累,土壤中根系吸收积累的Ni会被运输到小麦籽实等器官中.任艳军等(2016)的研究结果表明,孕穗期Ni主要吸收积累于根系(分配率占47.93%),穗中Ni分配率只占4.83%.进入成熟期,Ni主要积累分配于茎秆(分配率占32.04%)和籽实(分配率占22.28%)中,这与前人(Page et al., 2005)的研究结论一致.Ni的毒害效应造成成熟期籽实产量明显下降(表 3),成熟期籽实中Ni含量增幅均值(2022.12%)和Ni富集系数均值(0.15) 均高于孕穗期穗中相应指标大小,表明小麦孕穗期至成熟期阶段,小麦生殖器官籽粒中Ni吸收富集能力持续增加,从食用安全角度考虑,这是应该值得注意的问题.
4.2 Ni对小麦毒害临界值有关作物中重金属毒性临界值,一般采用达标临界值,即可食部位达食品卫生标准时的土壤重金属含量值.目前我国和国际上还缺乏有关Ni在粮食作物中的限量标准,但在本研究中,Ni在小麦籽粒中的含量随外源Ni的增加而增加.背景土壤条件下,小麦籽粒Ni含量为1.43 mg·kg-1,参照GB 2762-2012中油脂及其制品Ni的限量标准(1.0 mg·kg-1),小麦籽粒中Ni含量超过该项标准.但有报道表明,近10年我国粮食产区小麦中Ni的含量在0.2~47.5 mg·kg-1之间,算术平均值为15.10 mg·kg-1 (黄锦孙等,2012).本试验中,25和50 mg·kg-1处理,籽实Ni含量分别为7.39和8.05 mg·kg-1,明显低于全国粮食产区Ni算数平均值水平(15.10 mg·kg-1),而土壤环境质量标准中(GB15618—1995) 为保障农业生产,维护人体健康的土壤限制值为50 mg·kg-1;Ni添加量超过100 mg·kg-1以上时,100、200和400 mg·kg-1Ni处理,籽实中Ni含量分别为18.00、38.47和79.82 mg·kg-1(p < 0.01),均超过全国粮食产区Ni算数平均值水平(15.10 mg·kg-1).本研究提出,鉴于小麦作物籽实Ni的易吸收富集特性,对于粮食作物中Ni限量标准的制定,应予以高度重视.
由于谷物中Ni的食品卫生标准尚未制定,所以一般以作物减产5%~10%时的土壤全量Ni或土壤有效态Ni含量作为毒害临界值,减产幅度的选择主要是产量的数额,数额大者取低限,数额小者取高限(青长乐等,1992).本研究结果显示,外源Ni会降低小麦孕穗期茎秆、穗干质量和成熟期籽实千粒重和单区产量,土壤中添加Ni量达到100 mg·kg-1时,各项指标大小与对照比较均显著下降;方差分析结果均达到显著差异水平(p < 0.05).孕穗期和成熟期Ni毒害效应的突变点非常吻合,但由于成熟期籽实千粒重和单区产量在统计学上的数额差异,其减产幅度分别为7.29%和15.32%,小麦籽实产量降幅均值为11.30% (EC10).综合上述讨论食品卫生标准中小麦籽实中Ni含量的评价结论,确定本研究中Ni的毒害临界值约为Ni添加量为100 mg·kg-1处理下的土壤全Ni量139.08 mg·kg-1.据报道,作物受害时土壤中Ni的临界值大多在50~150 mg·kg-1范围内(张素霞等,1991).相关分析结果显示,土壤中不同形态Ni(HCl和DTPA提取)和根系、穗中Ni含量分别与小麦孕穗期茎秆和穗干质量呈显著负相关(p < 0.05, p < 0.01),同时与成熟期籽实千粒重和单区产量呈显著负相关(p < 0.05, p < 0.01).为了验证小麦发育前期外源Ni毒害效应对成熟期籽实产量的影响,小麦孕穗期茎秆和穗干质量与成熟期籽实千粒重和单区产量均表现出显著的正相关关系(p < 0.05),表明通过研究作物不同生育期毒害效应的指标变化,实现作物Ni毒害临界指标的中期诊断是可行的.本研究提出,小麦孕穗期相关组织器官毒害效应的突变点对应的指标(统计学差异达显著水平及以上)可作为小麦Ni毒害临界指标.根据文献报道提出的作物植株体内Ni、土壤有效态Ni (DTPA)、根系Ni含量均可作为小麦作物的毒害临界指标(青长乐等,1992;杨定清等,1994;马建军等,2006).除土壤全Ni毒害临界值外,土壤有效态Ni (DTPA提取)含量(21.59 mg·kg-1)、小麦穗Ni含量(19.17 mg·kg-1,与文献(青长乐等,1992)毒害临界值>10 mg·kg-1吻合)、小麦根系Ni含量(318.49 mg·kg-1)均可作为小麦外源Ni毒害效应的临界值.
5 结论(Conclusions)1) 外源Ni会增加小麦孕穗期株高和叶片干质量,抑制茎秆和穗干物质积累,降低成熟期小麦籽实千粒重和单区产量;Ni投加量为25~50 mg·kg-1时,毒害效应指标变化不明显,Ni投加量为100 mg·kg-1及以上时,毒害效应指标抑制作用明显,Ni是造成小麦毒害效应的主要因素.
2) 外源Ni会增加小麦孕穗期和成熟期各器官中Ni的含量,小麦孕穗期各器官中Ni主要吸收积累于根系,地上部各器官中,穗中Ni含量和富集系数均高于其它器官,孕穗期至成熟期阶段小麦植株中Ni易向生殖器官籽粒中吸收和富集.
3) 小麦孕穗期毒害效应与成熟期籽实产量之间存在显著正相关关系,且毒害效应突变点一致,以小麦孕穗期Ni毒害效应突变点作为临界指标表征外源Ni小麦成熟期籽实产量抑制效应是可行的,提出小麦作物Ni毒害中期诊断指标:土壤全Ni含量为139.08 mg·kg-1,土壤有效态Ni(DTPA提取)含量为21.59 mg·kg-1,小麦穗Ni含量为19.17 mg·kg-1,小麦根系Ni含量为318.49 mg·kg-1.
| [${referVo.labelOrder}] | 白英, 刘铮, 刘占刚, 等. 2014. 外源Ni在干旱区绿洲土壤-小麦系统中的迁移及其生物有效性[J]. 环境科学学报, 2014, 34(7): 1801–1807. |
| [${referVo.labelOrder}] | Eskew D L, Welch R M, Cary E E. 1983. Nickel:an essential micronutrient for legumes and possibly all higher plants[J]. Science, 222(4642): 621–623. |
| [${referVo.labelOrder}] | Gajewska E, Sklodowska M, Slaba M, et al. 2006. Effect of nickel on antioxidative enzyme activities, proline and chlorophyll contents in wheat shoots[J]. Biologia Plantarum, 50(4): 653–659. DOI:10.1007/s10535-006-0102-5 |
| [${referVo.labelOrder}] | 黄锦孙, 韦东普, 郭雪雁, 等. 2012. 田间土壤外源铜镍在小麦中的累积及其毒害研究[J]. 环境科学, 2012, 33(4): 1369–1375. |
| [${referVo.labelOrder}] | 蒋德富, 陈帆. 1987. 镍、钒对春小麦生长发育及产量影响的研究[J]. 环境科学, 1987, 8(4): 41–44. |
| [${referVo.labelOrder}] | 刘国栋. 2001. 植物营养元素-Ni[J]. 植物营养与肥料学报, 2001, 7(1): 103–108. DOI:10.11674/zwyf.2001.0115 |
| [${referVo.labelOrder}] | 李雪, 高天鹏, 吕亚娟, 等. 2015. 外源水杨酸对小麦幼苗镍毒害的缓解效应[J]. 麦类作物学报, 2015, 35(3): 402–407. DOI:10.7606/j.issn.1009-1041.2015.03.017 |
| [${referVo.labelOrder}] | Molas J. 1998. Changes in morphological and anatomical structure of cabbage (Brassica oleracea L.) outer leaves and in ultrastructure of their chloroplasts caused by an in vitro excess of nickel[J]. Photosynthetica, 34(4): 513–522. |
| [${referVo.labelOrder}] | 马建军, 于凤鸣, 朱京涛, 等. 2006. 4种浸提剂对土壤有效态镍提取条件的比较与选择[J]. 农业环境科学学报, 2006, 25(增刊): 746–750. |
| [${referVo.labelOrder}] | Patrick H B, Ross M W, Cary E E, et al. 1987. Beneficial effects of nickel on plant growth[J]. Journal of Plant Nutrition, 10: 2125–2135. DOI:10.1080/01904168709363763 |
| [${referVo.labelOrder}] | Page V, Feller U. 2005. Selective transport of zinc, manganese, nickel, cobalt and cadmium in the root system and transfer to the leaves in young wheat plants[J]. Annals of Botany, 96(3): 425–434. DOI:10.1093/aob/mci189 |
| [${referVo.labelOrder}] | 青长乐, 牟树森, 蒲富永, 等. 1992. 论土壤重金属毒性临界值[J]. 农业环境保护, 1992, 11(2): 51–56. |
| [${referVo.labelOrder}] | 任艳军, 任学军, 彭飞, 马建军. 2017. 利用小麦三叶期毒害效应指标标征成熟期籽实产量Ni毒害的探讨[J]. 环境科学学报, 2017. DOI:10.13671/j.hjkxxb.2017.0051 |
| [${referVo.labelOrder}] | 任艳军, 任学军, 彭飞, 马建军. 2016. 春小麦不同生育期各器官主要矿质元素吸收和分配的动态变化[J]. 河北科技师范学院学报, 2016, 30(4): 38–45. |
| [${referVo.labelOrder}] | Ramos L, Hernandez L M, Gonzalez M J. 1994. Sequential fractionation of copper lead, cadmium and zinc in soils from or near Donana National park[J]. Journal of Environmental Quality, 23(1): 50–57. |
| [${referVo.labelOrder}] | Seregin I V, Kozhevnikova A D, Davydova M A, et al. 2007. Role of root and shoot tissues of excluders and hyperaccumulators in nickel transport and accumulation[J]. Doklady Biological Sciences, 415(1): 295–297. DOI:10.1134/S0012496607040138 |
| [${referVo.labelOrder}] | 王夔. 1992. 生命科学中的微量元素(下卷)[M]. 北京: 中国计量出版社: 482. |
| [${referVo.labelOrder}] | Wang P X, Qu E F, Li Z B, et al. 1997. Fractions and availability of nickel in loessial soil amended with sewage or sewage sludge[J]. Journal of Environmental Quality, 26(3): 795–801. |
| [${referVo.labelOrder}] | 杨红超, 王素平. 2011. 重金属镍胁迫对小麦种子萌发及幼苗生长的影响[J]. 种子, 2011, 30(12): 18–20. DOI:10.3969/j.issn.1001-4705.2011.12.006 |
| [${referVo.labelOrder}] | 杨林书, 吴娇掀, 王宏康. 1996. 用小麦幼苗毒性指标标征土壤Cd污染的研究[J]. 农业环境保护, 1996, 15(2): 81–85. |
| [${referVo.labelOrder}] | 杨定清, 傅绍清, 青长乐. 1994. 镍的作物效应及临界值研究[J]. 四川环境, 1994, 13(1): 19–23. |
| [${referVo.labelOrder}] | 朱京涛, 马建军, 杜彬, 等. 2008. 不同种类Ni2+盐浸种对作物种子活力的影响[J]. 安徽农业科学, 2008, 36(10): 3973–3974. DOI:10.3969/j.issn.0517-6611.2008.10.020 |
| [${referVo.labelOrder}] | 张素霞, 王宏康. 1991. 污泥施肥时重金属镍对农作物的毒害研究[J]. 环境科学学报, 1991, 11(1): 71–78. |
| [${referVo.labelOrder}] | 中华人民共和国卫生部. 2012. GB 2762-2012食品安全国家标准食品中污染物限量[S]. 北京: 中国标准出版社 |