 2017, Vol. 37
2017, Vol. 37



2. 福建农林大学林学院, 福州 350002
2. College of Forestry, Fujian Agriculture and Forestry University, Fuzhou 350002
膜生物反应器(MBR)是一种膜分离技术与活性污泥法相结合的新型处理工艺, 与传统污水处理技术相比, 具有分离效率高、占地面积小、出水水质好、污泥产量低等优点, 然而在工程实践过程中, 膜污染问题却一直限制着该技术的推广与应用(Tan et al., 2015) .
膜生物污染的本质是污泥中微生物在膜表面及孔隙内部形成的生物膜, 而在此过程中微生物与膜材料间的吸附作用扮演着重要角色.已有的研究表明, 微生物在固体表面间的吸附是一个复杂的热力学过程, 与吸附双方的表面物化性质密切相关, 并伴随着二者界面交互作用自由能的变化.Li等(2004) 在研究8株细菌在不同类型玻璃表面的吸附行为时发现, 相较于普通玻璃, 微生物菌体更倾向吸附在金属氧化物修饰的玻璃表面, 而进一步研究则发现, 这些金属氧化物的存在提升了微生物菌体与玻璃界面的吸附自由能;Lopes等(2005) 以Desulfovibrio desulfuricans菌为模型, 比较了其在金属镍、不锈钢及有机玻璃3种介质表面的吸附差异, 并发现不同材料在亲/疏水性、荷电性、粗糙度等方面的差异是影响菌体吸附行为的主要因素;Gillett等(2015) 则通过激光改变菌体表面的性质后发现, E. coli在其他物质表面的附着与分布发生了显著变化.在膜污染方面, Kang等(2004) 以酵母菌及葱伯克霍尔德菌为模型, 结合显微图像技术与界面作用力计算, 对其在错流过滤过程中的膜吸附特征进行了研究, 发现过滤中的渗透力与双层静电力是这些微生物膜吸附的主要作用力;Noeon等(2005) 则以黄杆菌属所产的溶解性微生物产物(SMP)为例, 通过经典热力学的扩展DLVO (XDLVO)理论研究了其在4种纳滤膜上的吸附特征, 结果发现, 由于界面间存在较高的Lewis酸碱作用力, 导致不同纳滤膜均对该种SMP产生不同程度的排斥作用;赵飞等(2015) 以铜绿假单胞菌(PA)为模式菌株, 分析了Ca2+浓度、海藻酸钠、牛血清白蛋白和腐殖酸对纳滤膜生物污染的影响, 结果表明, 不同有机物和Ca2+浓度对吸附系统的范德华作用能影响不大, 而对疏水作用能则有着显著影响.目前, 有关膜生物污染中微生物吸附行为的研究相对较少, 仅有的一些研究也主要集中在纳滤和超滤膜方面, 且多是以一些模拟物为研究对象, 而真正涉及MBR膜污染层微生物膜吸附行为的则罕见报道.
基于此, 本文以2株分离至MBR膜污染层的微生物Aeromonas caviae N25与Aeromonas veronii M4为研究对象, 结合热力学方法及XDLVO理论对其与聚丙烯(PP)微滤膜间的界面交互作用进行研究, 分析这2种细菌在PP膜界面的吸附机制.其结果对于完善膜生物污染相关理论、深入理解膜污染的本质及制定合理有效的膜污染控制策略具有重要的理论价值和实际指导意义.
2 材料与方法(Materials and methods) 2.1 主要试剂Na2HPO4、NaH2PO4、NaHCO3、Na2CO3、HCl、酵母提取粉、胰蛋白胨、琼脂粉、NaCl、甘油、多聚甲苯购自国药集团, 对苯二胺购自阿拉丁公司, 荧光染料SYBR Green I购自Solarbio公司.
2.2 实验耗材聚丙烯膜购自浙江能大过滤设备有限公司, 直径100 mm, 孔径0.22 μm;六孔板购自美国Corning.
2.3 菌株Aeromonas caviae N25、Aeromonas veronii M4为本实验室收藏, 为前期筛选自一台处理生活废水MBR膜污染层的2株优势菌.而Aeromonas菌属也已被证明在MBR 膜污染层中大量存在, 并在膜污染过程中起着重要的作用(Harshad et al., 2014; Zhao et al., 2015) .
2.4 细菌悬液的制备将A.caviae N25与A.veronii M4在LB培养基中培养的种子液, 按1%的比例分别接种于LB液体培养基中, 振荡培养至稳定阶段, 4 ℃、5000 g 条件下离心10 min, 弃上清液, 采用无菌水对菌体进行清洗离心2次后, 重悬于无菌水中, 浓度调为2×108 CFU·mL-1(A.caviae N25的OD600=1.2, A.veronii M4的OD600=0.9) .
2.5 菌株、膜性质的分析 2.5.1 菌体大小采用荧光显微镜(BK-FL4, 重庆奥特)观察、拍照, 使用ISCapture软件测量菌体半径(Zheng et al., 2014) .
2.5.2 菌体、膜表面的接触角菌体表面接触角的测定采用Brant等(2002) 的方法, 选用3种标准液体(水、甘油、二碘甲烷), 通过接触角测定仪(DSA100, 德国Kruss)分别测定细菌与PP膜表面的接触角.
2.5.3 菌体、膜表面的Zeta电位将收集得到的菌体, 用无菌的1 mmol·L-1 KCl 溶液制成浓度为2×108 CFU·mL-1的菌悬液, 利用Zeta电位仪(ZS90, 马尔文)测定菌悬液的电位.PP膜表面Zeta电位则采用流动电位仪(Surpass, 安东帕)测定(Pradeepa et al., 2016) .
2.5.4 菌体疏水性的分析以正辛烷为有机相溶剂, 采用烃类化合物吸附法(BATH)测定菌体表面疏水性 (Heilmann et al., 1996) .
2.5.5 细菌运动性的研究分别采用TYG培养基、NA培养基与LB培养基3种培养基(Pang et al., 2005), 对2株细菌的泳动(0.3%琼脂)、蠕动(0.5%琼脂)与蹭动(1%琼脂)等运动特征进行评价.
2.6 细菌的静态吸附实验将PP膜置于六孔板内, 分别向孔内添加2种细菌的菌悬液(2×108 CFU·mL-1), 7 mL·孔-1;然后在30 ℃、100 r·min-1条件下培养, 并于不同阶段取样, 用无菌水轻轻冲去膜表面浮菌, 4%多聚甲苯固定12 h;最后室温晾干.
2.7 静态吸附实验的荧光显微镜分析将静态吸附后的PP膜用1:1000的SYBR Green I工作液染色15 min, 在激发波长460 nm条件下, 利用荧光显微镜(BK-FL4, 重庆奥特)观察, 并用ISCapture软件对吸附在膜表面的细菌进行计数.
2.8 原子力显微镜(AFM)分析当静态吸附实验结束后, 利用AFM (5500, 美国 Agient)观察2株细菌在PP膜上的吸附特征, 样品制作方法同2.6节.采用氮化硅探针, 获得扫描10 μm×10 μm区域, 通过计算吸附前后膜表面的高低位差值、平均粗糙度(Rq)等指标分析细菌在膜表面的吸附情况(Sylwia et al., 2014) .
2.9 扫描电镜(SEM)分析当静态吸附实验结束后, 采用扫描电镜(Nova NanoSEM 230, 美国FEI)对2种细菌在PP膜上的吸附特征进行观察(Chen et al., 2015), 样品制作方法同2.6节.
2.10 热力学参数固体表面能、液体表面能及二者之间接触角的关系见式(1)和(2) (Brant et al., 2002) .
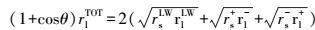
|
(1) |
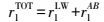
|
(2) |
式中, θ为固体与液体表面的接触角(°), rlTOT为液体的表面能(mJ·m-2), rlLW为范德华分量(mJ·m-2), rlAB为Lewis酸碱作用分量(mJ·m-2), rl+、rl-分别为Lewis酸分量(mJ·m-2)和Lewis碱分量(mJ·m-2), 下标s、l分别表示固体、液体.
固体物质界面能ΔGsws(mJ·m-2)与Lewis酸分量r+和Lewis碱分量r-可表示为式(3) ~(5) (Van Oss et al., 1995) .当ΔGsws>0 mJ·m-2时, 该物质是亲水的;反之, 则是疏水的.其中, w表示水.
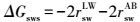
|
(3) |
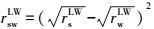
|
(4) |
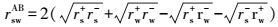
|
(5) |
XDLVO理论微生物与膜表面的界面能见式(6) ~(9) (Brant et al., 2002) .膜-微生物体系界面自由能的范德华与Lewis酸碱分量可以表示为式(10) ~(11) (Brant et al., 2002) .
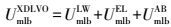
|
(6) |
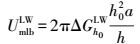
|
(7) |
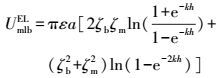
|
(8) |
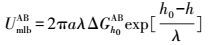
|
(9) |
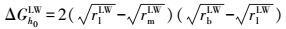
|
(10) |
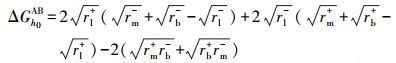
|
(11) |
式中, UmlbXDLVO为膜-微生物表面的总界面能(kT);UmlbLW为膜-微生物体系界面能的范德华分量(kT);UmlbEL为膜-微生物体系界面能的静电力分量(kT);UmlbAB为膜-微生物体系界面能的Lewis酸碱分量(kT);a为微生物菌体半径(μm);h0为微生物与膜表面的最小作用距离, 取值0.158 nm (Bhattacharjee et al., 1996) ;h为微生物与膜表面的作用距离(nm);λ为衰减长度, 取值为0.6 nm (Bhattacharjee et al., 1996) ;ε为分散介质的绝对介电常数(F·m-1);k为Debye长度的倒数, 常温下取0.104 nm;ζm与ζb分别代表膜与微生物表面的Zeta电位(mV).
2.12 吸附动力学模型2株细菌在PP膜表面的吸附反应动力学采用准一级动力学方程(式(12), Lagergren et al., 1989) 、准二级动力学方程(式(13), Skodras et al., 2008) 进行描述.
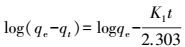
|
(12) |
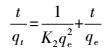
|
(13) |
式中, K1(min-1)、K2(CFU·cm-2·min-1)分别为准一级、准二级反应吸附速率;qe、qt分别为吸附平衡、吸附时间为t时的单位面积PP膜吸附菌体的量(CFU·cm-2).
3 结果与讨论(Results and discussion) 3.1 微生物的疏水性分析A.caviae N25和A.veronii M4的疏水性特征如图 1所示.从图中可知, 2株菌的疏水性能具有相似的变化特征, 均随着有机相正辛烷添加量的增加呈现出先上升后下降的变化趋势.在初始阶段, 2株菌的疏水率相对较低, 这可能是由于菌体浓度过大, 超出正辛烷的吸附能力, 致使一部分细菌滞留在水相所造成的.但当正辛烷-水相比大于6:5时, A.caviae N25与A.veronii M4的疏水率开始下降, 而这可能是过量的有机相对细胞活性产生抑制作用所造成的.然而, 尽管2株菌具有类似的变化趋势, 但二者的疏水性能具有显著差异, 在相同正辛烷体积条件下, A.veronii M4的疏水性能要明显高于A.caviae N25, 这可能是2种微生物菌体外壁蛋白质与脂多糖的比例不同所致(盛下放等, 2004) .
 |
| 图 1 A.caviae N25与A.veronii M4菌株的表面疏水性特征 Fig. 1 Cell-surface hydrophobic properties of A.caviae N25 and A.veronii M4 |
分别采用TYG、NA、LB 3种培养基, 对A.caviae N25和A.veronii M4的运动性能进行评测, 结果如表 1和图 2所示.在泳动方面, A.veronii M4菌在TYG与NA培养基上具有较好的泳动性能, 但在LB培养基上却受到了抑制, 而A.caviae N25则恰好相反;在蠕动方面, A.veronii M4菌在3种培养基中均表现出不同程度的蠕动性能, 特别是在TYG培养基上其拓展面积达到了0.8 cm2, 而A.caviae N25则只在TYG和LB培养基中具有较好的蠕动性能;另外, A.caviae N25与A veronii M4两株菌在3种培养基上均表现出较为明显的蹭动性能.综上所述, A.caviae N25菌株具有较好的蠕动与蹭动性能, 而A.veronii M4则在泳动与蠕动、蹭动性能方面较强.已有的研究表明, 微生物的运动性能与其在固体界面的粘附密切相关(Hall-Stoodley et al., 2002), 由于A.veronii M4具有相对较好的运动性能, 推断该菌株在非生物介质表面的粘附能力也应更强.
 |
| 图 2 不同培养基对A.caviae N25(a, b, c)、A.veronii M4(d, e, f)的运动能力的影响 Fig. 2 Effect of different media on the motility of bacterial isolates |
| 表 1 不同培养基对A.caviae N25、A.veronii M4运动能力的影响 Table 1 Effect of different media on the motility of bacterial isolates |

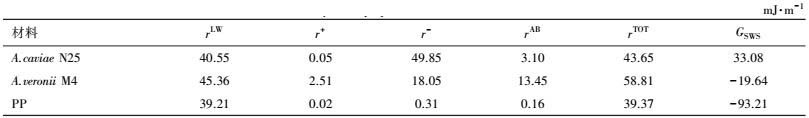
A.caviae N25、A.veronii M4与PP膜材料的表面热力学性质如表 2和表 3所示.由表 3可知, 3种标准液体在PP膜表面的接触角均明显高于A.caviae N25和A.veronii M4, 因此, PP膜表面具有明显的疏水性能.A.caviae N25、A.veronii M4菌株和PP膜的表面Zeta电位均为负值, 即3种物质表面均带有负电荷.已有的研究发现(Sheng et al., 2007), 微生物菌体表面之所以带有负电荷, 这是细胞表面的阴离子所致.并且3种物质表面的Zeta电位值与θW值呈正相关, 即Zeta电位值越大, θW越大, 该结论也与赵应许等(2014) 的研究结果相一致.另外, 由表 2可知, 3种物质中PP膜的GSWS绝对值最大, 因此, 其表面疏水性更强(Nguyen et al., 2011), 这也与前面的疏水性及接触角分析相一致.
| 表 2 A.caviae N25和A.veronii M4与PP膜材料的表面热学性质 Table 2 Surface thermodynamic properties of different bacteria and PP membrane |
| 表 3 A.caviae N25和A.veronii M4的表面性质 Table 3 Surface properties of A.caviae N25 and A.veronii M4 |
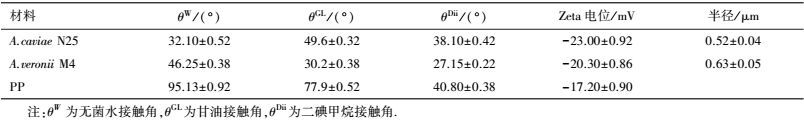
另外, 2株细菌及PP膜表面的rLW值相差并不大, 均为39~40 mJ·m-2, 但三者的r-值均大于r+值, 表现出较强的供电子能力特征, 而该结果也与Brant等(2002)和Kim等(2007) 的研究报道相一致.此外, 由于A.caviae N25和A.veronii M4菌株膜表面的r-比PP膜的r-大2个数量级, 使得PP膜的rAB值远小于A.caviae N25和A.veronii M4的rAB值;而尽管A. veronii M4的r-明显小于A. caviae N25的r-, 但因具有较高的r+, 使得A. veronii M4具有较高的rAB.
3.4 微生物与膜系统间的界面能分析图 3所示为A.caviae N25-PP膜系统和A.veronii M4-PP膜系统之间的界面自由能变化特征.从图中可知, 在2种吸附体系内都是Lewis酸碱分量UmlbAB绝对值最大, 静电力分量UmlbEL最小, 说明Lewis酸碱分量UmlbAB在2株菌的膜吸附进程中占据主导地位;吸附体系内的UmlbLW和UmlbAB均为负值, 表明这2种界面能分量会促进细菌在膜表面上的吸附;而UmlbEL为正值, 说明该界面能分量起着吸附排斥作用, 这可能是由于膜与菌体表面均带负电荷, 电性相同所致.A.caviae N25-PP膜系统中UmlbEL出现了负值, 而Brant等(2002) 在膜与胶体相互作用的实验过程中也发现具有相同电荷属性的物质表面之间发生相互吸引作用, 该结果可能是由于物质表面电荷密度差值较大所造成的.另外, 微生物与膜表面之间的界面能受距离的影响较大, 尤其是Lewis酸碱分量, 当菌体与膜之间的距离小于4 nm时, UmlbAB绝对值迅速增大, 而作为长程作用力的静电力分量UmlbEL, 变化较弱.
 |
| 图 3 不同微生物-膜系统界面能随距离的变化 (a.A.caviae N25, b.A.veronii M4, c.总界面能) Fig. 3 Interaction energy components variation of the two bacterium-PP membrane systems as a function of separation distance (a. A.caviae N25, b. A.veronii M4, c. The total interaction energy) |
从图 3c可知, A.caviae N25、A.veronii M4与PP膜间的界面总作用能均为负值, 表明2种微生物在PP膜表面的吸附行为是一个自发进行的过程(李坤权等, 2010) ;另外, 由于A.veronii M4-PP膜间的界面作用能(绝对值)明显大于A.caviae N25-PP膜系统, 则理论上表明A.veronii M4菌株更容易克服与PP膜之间的作用能垒, 吸附在PP膜表面.
3.5 微生物对PP膜的吸附行为A.caviae N25和A.veronii M4在PP膜表面的静态吸附结果如图 4和图 5所示.从图中可知, 2种细菌在PP膜表面呈现出不同的吸附走势, 对于A.caviae N25而言, 其在最初的20 min内就快速完成了吸附, 后续时间吸附量变化不大, 而A.veronii M4则在经过60 min后, 其吸附才逐渐变得平缓并趋于稳定;80 min时, 2株菌的吸附均达到平衡阶段;另外, 与A.caviae N25相比, 尽管A.veronii M4需要较长的吸附平衡时间, 但在相同时间内A.veronii M4在PP膜上的吸附量要明显高于A.caviae N25.这也进一步表明A.veronii M4对PP膜具有较强的吸附能力, 此结论也与前面系统界面能的分析结果相一致, 表明微生物菌体对固体界面的吸附能力与二者的系统界面作用能密切相关.相关研究表明, 微生物与非生物介质表面性质及二者之间界面作用的不同会导致微生物与非生物介质之间吸附行为的差异(Palmer et al., 2007) .本研究中, A.caviae N25和A.veronii M4的疏水性、电荷性、电位等表面性质差异明显, 这些性质的不同使得不同微生物与膜之间呈现不同的界面作用, 因此, 造成了2种微生物吸附PP膜表面不同的吸附特征.上述微生物-膜系统之间界面能的分析结果中A.veronii M4-PP膜间的界面能(绝对值)大于A.caviae N25-PP膜系统, 即A.veronii M4菌株更易造成膜污染, 该结论也与已有的一些研究报道相一致(Bayoudha et al., 2009) .
 |
| 图 4 两株菌对PP膜吸附的SYBR Green染色结果 Fig. 4 Fluroesence staining images of PP membrane adsorbed by two strains |
 |
| 图 5 A.caviae N25和A.veronii M4对PP膜的吸附量 Fig. 5 Adsorption capacity of A.caviae N25 and A.veronii M4 to PP membrane |
采用AFM对经菌体吸附后的PP膜进行图像分析, 结果见图 6.结果表明, 在吸附发生前后, PP膜表面的粗糙度、高低差都发生了明显的变化.静态吸附之前, PP膜表面相对比较平整, 高低差起伏不大(高低差为0.425 μm), 经A.caviae N25与A.veronii M4吸附后, 其表面高低差明显增大, 分别达到了1.6 μm与0.74 μm;另外, 由表 4可知, 在A.caviae N25与A.veronii M4吸附过后, PP膜表面粗糙度由原来的22.0 nm分别达到了73.9 nm和48.5 nm, 前后变化明显.由前面的吸附数据可知, 相同时间内, A.veronii M4在PP膜上的吸附量要明显高于A.caviae N25, 伴随吸附数量的增加, 也使得该菌株在膜表面的分布相对均匀, 进而导致了相对较低的表面高低差及粗糙度.该研究结果与Xu等(2002) 在微生物腐蚀的研究中的结果类似, 被菌体吸附之后的PP膜表面性质发生了明显的变化(如粗糙度).
 |
| 图 6 A.caviae N25与A.veronii M4吸附前后PP膜的AFM图谱 (a. 空白膜, b. A.caviae N25吸附后的膜, c. A. veronii M4吸附后的膜) Fig. 6 AFM images of PP membrane before and after adsorbed by A. caviae N25 and A. veronii M4 |
| 表 4 A.caviae N25与A.veronii M4吸附前后PP膜的Rq值 Table 4 Rq value of PP membrane before and after adsorbed by A.caviae N25 and A.veronii M4 |

利用SEM对2株菌在PP膜上的吸附特征进行比较(图 7) .从图中可知, A.caviae N25与A.veronii M4的细胞形状均为长杆状, 并随机吸附在PP膜的纤维缝隙与材料表面;相较于A.caviae N25, 菌株A.veronii M4在PP膜上表现出明显的吸附优势, 且分布的相对比较均匀, 有效地减少了一些缝隙的存在, 进而导致其具有较小的表面粗糙度, 这也进一步支持了AFM的检测结果.
 |
| 图 7 A.caviae N25(a)与A veronii M4(b)吸附过后PP膜的SEM图谱 Fig. 7 SEM images of PP membrane after adsorption by A.caviae N25(a) and A.veronii M4(b) |
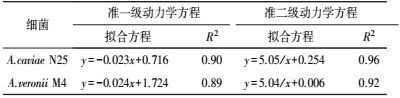
采用准一级、准二级动力学模型对2株细菌的膜吸附行为进行拟合分析, 结果如图 8与表 5所示.从中可知, 准二级动力学模型的R2高于相对应的准一级动力学模型, 表明准二级动力学模型更适合描述2株菌在PP膜上的吸附行为.而一些相关研究也发现, 准一级动力学模型更适合描述吸附初期的动力学过程, 并不适合描述整个吸附过程(李坤权等, 2010) .因此, A.caviae N25与A.veronii M4两株菌在PP膜表面的吸附过程可采用准二级动力学模型进行描述.
 |
| 图 8 两株微生物对 PP膜吸附的动力学方程拟合 (a.准一级动力学, b.准二级动力学) Fig. 8 The pseudo-first-order (a) and pseudo-second-order (b) kinetics fit of PP membrane adsorbed by two strains |
| 表 5 两株微生物对PP膜吸附的动力学方程参数 Table 5 Kinetics parameters of PP membrane adsorbed by two strains |
1) A.veronii M4与A.caviae N25在正辛烷中的疏水性均呈现先增加后减少的趋势, 但相同条件下, A.veronii M4的疏水性能相对较高, 而A.veronii M4的运动性能也要强于A.caviae N25.
2) 2株细菌与PP膜的r-值均大于r+, 且Zeta电位都为负值, 说明其表面带有负电荷, 并具有较强的电子供体特征, 而A.veronii M4与PP膜的GSWS为负值, 则代表其具疏水性特征, 而A.caviae N25的GSWS为正值, 表现为亲水性特征.
3) 在2株细菌与PP膜的界面自由能中, UmlbLW和均UmlbAB为负值, 对细菌的吸附起促进作用, 而UmlbEL为正值, 则对细菌吸附起抑制作用;另外, Lewis酸碱分量UmlbAB在3种界面能中绝对值最大, 因此, 占据主导地位;2株细菌与PP膜的总界面作用能均为负值, 意味着其在膜上的吸附是一个主动过程, 而A.veronii M4与PP膜间具有较大的总界面作用能, 表明该菌株具有相对较强的吸附能力.
4) 通过荧光染色、AFM及SEM分析2株菌株在PP膜的吸附特征发现, A.veronii M4在膜上的吸附量要明显高于A.caviae N25;而吸附动力学的拟合结果发现, 2株菌的吸附过程均符合准二级动力学模型.
| [${referVo.labelOrder}] | Bayoudha S, Othmaneb A, Morac L, et al. 2009. Assessing bacterial adhesion using DLVO and XDLVO theories and the jet impingement technique[J]. Colloids and Surfaces B: Biointerfaces, 73(1): 1–9. DOI:10.1016/j.colsurfb.2009.04.030 |
| [${referVo.labelOrder}] | Bhattacharjee S, Sharma A, Bhattacharya P K. 1996. Estimation and influence of long range solute:membrane interactions in ultrafiltration[J]. Industrial and Engineering Chemistry Research, 35(9): 3108–3121. DOI:10.1021/ie9507843 |
| [${referVo.labelOrder}] | Brant J A, Childress A E. 2002. Assessing short-range membrane-colloid interactions using surface energetics[J]. Journal of Membrane Science, 203(1/2): 257–273. |
| [${referVo.labelOrder}] | Chen P Y, Cui L, Zhang K S. 2015. Surface-enhanced Raman spectroscopy monitoring the development of dual-species biofouling on membrane surfaces[J]. Journal of Membrane Science, 473: 36–44. DOI:10.1016/j.memsci.2014.09.007 |
| [${referVo.labelOrder}] | Gillett A R,Waugh D G,Lawrence J.2015.Influencing the attachment of bacteria through laser surface engineering[C].The 34th International Congress on Applications of Lasers and Electro-Optics (ICALEO 2015).Atlanta,GA,USA:18-22 |
| [${referVo.labelOrder}] | Hall-Stoodley L, Stoodley P. 2002. Developmental regulation of microbial biofilms[J]. Current Opinion in Biotechnology, 13(3): 228–233. DOI:10.1016/S0958-1669(02)00318-X |
| [${referVo.labelOrder}] | Harshad L, Diby P, Ji H K. 2014. Isolation and molecular characterization of biofouling bacteria and profiling of quorum sensing signal molecules from membrane bioreactor activated sludge[J]. International Journal of Molecular Sciences, 15(2): 259–264. |
| [${referVo.labelOrder}] | Heilmann C, Gerke C, Perdreau Remington F, et al. 1996. Characterization of Tn917 insertion mutants of Staphylococcus epidermidis affected in biofilm formation[J]. Infection and Immunity, 64(1): 277–282. |
| [${referVo.labelOrder}] | Kang S T, Subramani A, Hoek E M V, et al. 2004. Direct observation of biofouling in cross-flow microfiltration:mechanisms of deposition and release[J]. Journal of Membrane Science, 244: 151–165. DOI:10.1016/j.memsci.2004.07.011 |
| [${referVo.labelOrder}] | Kim S, Hoek E M V. 2007. Interactions controlling biopolymer fouling of reverse osmosis membranes[J]. Desalination, 202(1/3): 333–342. |
| [${referVo.labelOrder}] | Lagergren S. 1989. Zur theorie der sogenannten adsorption gelöster stoffe[J]. Handlingar, 24(4): 1–39. |
| [${referVo.labelOrder}] | Lopes F A, Morin P, Oliveira R, et al. 2005. The influence of nickel on the adhesion ability of Desulfovibrio desulfuricans[J]. Colloids and Surfaces B:Biointerfaces, 46(2): 127–133. DOI:10.1016/j.colsurfb.2005.07.020 |
| [${referVo.labelOrder}] | 李坤权, 郑正, 蒋剑春, 等. 2010. 铅在棉秆基活性炭上的吸附动力学与热力学[J]. 环境科学, 2010, 31(5): 1402–1408. |
| [${referVo.labelOrder}] | Nguyen V T, Chia T W R, Turner M S, et al. 2011. Quantification of acid-base interactions based on contact angle measurement allows XDLÜO predictions to attachment of Campylobacter jejuni but not Salmonella[J]. Journal of Microbiological Methods, 86(1): 89–96. DOI:10.1016/j.mimet.2011.04.005 |
| [${referVo.labelOrder}] | Noeon P, Boksoon K, Kim In S, et al. 2005. Biofouling potential of various NF membranes with respect to bacteria and their soluble microbial products (SMP):Characterizations,flux decline,and transport parameters[J]. Journal of Membrane Science, 258: 43–54. DOI:10.1016/j.memsci.2005.02.025 |
| [${referVo.labelOrder}] | Palmer J, Flint S, Brooks J. 2007. Bacterial cell attachment,the beginning of a biofilm[J]. Journal of Industrial Microbiology & Biotechnology, 34(9): 577–588. |
| [${referVo.labelOrder}] | Pang C M, Hong P, Guo H, et al. 2005. Biofilm formation characteristics of bacterial isolates retrieved from a reverse osmosis membrane[J]. Environmental Science & Technology, 39(19): 7541–7550. |
| [${referVo.labelOrder}] | Pradeepa, Akshay D S, Koshi M, et al. 2016. Multidrug resistant pathogenic bacterial biofilm inhibition by lactobacillus plantarum exopolysaccharide[J]. Bioactive Carbohydrates and Dietary Fibre, 8(1): 7–14. DOI:10.1016/j.bcdf.2016.06.002 |
| [${referVo.labelOrder}] | Skodras G, Diamantopoulou I, Pantoleontos G, et al. 2008. Kinetic studies of lemental mercury adsorption in activated carbon fixed bed reactor[J]. Journal of Hazardous Materials, 158(1): 1–13. DOI:10.1016/j.jhazmat.2008.01.073 |
| [${referVo.labelOrder}] | 盛下放, 何琳燕, 龚建勋. 2004. 二株假单胞菌的疏水性及其对菲的降解能力[J]. 环境科学学报, 2004, 24(5): 942–944. |
| [${referVo.labelOrder}] | Sheng X X, Ting Y P, Pehkonen S O J. 2007. Force measurements of bacterial adhesion on metals using a cell probe atomic forcemicroscope[J]. Journal of colloid and interface science, 310(2): 661–669. DOI:10.1016/j.jcis.2007.01.084 |
| [${referVo.labelOrder}] | Sylwia M, Dominika D, Aleksander O, et al. 2014. Microscopic studies on TiO2 fouling of MF/UF polyethersulfone membranes in a photocatalytic membrane reactor[J]. Journal of Membrane Science, 470: 356–368. DOI:10.1016/j.memsci.2014.07.049 |
| [${referVo.labelOrder}] | Tan J M, Qiu G l, Ting Y P. 2015. Osmotic membrane bioreactor for municipal wastewater treatment and the effects of silver nanoparticles on system performance[J]. Journal of Cleaner Production, 88: 146–151. DOI:10.1016/j.jclepro.2014.03.037 |
| [${referVo.labelOrder}] | Van Oss C J, Giese R. 1995. The hydrophilicity and hydrophobicity of clay minerals[J]. Clays and Clay Minerals, 43(4): 474–477. DOI:10.1346/CCMN |
| [${referVo.labelOrder}] | Xu L C, Chan K Y, Fang H H P. 2002. Application of atomic force microscopy in the study of microbiologically influenced corrosion[J]. Materials Characterization, 48(2/3): 195–203. |
| [${referVo.labelOrder}] | 赵飞, 许柯, 任洪强, 等. 2015. XDLÜO 理论解析有机物和钙离子对纳滤膜生物污染的影响[J]. 中国环境科学, 2015, 35(12): 3602–3611. DOI:10.3969/j.issn.1000-6923.2015.12.010 |
| [${referVo.labelOrder}] | Zhao X, Wang X C, Chen Z L, et al. 2015. Microbial community structure and pharmaceuticals and personal care products removal in a membrane bioreactor seeded with aerobic granular sludge[J]. Applied Microbiology and Biotechnology, 99(1): 425–433. DOI:10.1007/s00253-014-5984-0 |
| [${referVo.labelOrder}] | 赵应许, 纵瑞强, 高欣玉, 等. 2014. XDLÜO理论解析不同离子条件下海藻酸钠微滤膜污染[J]. 环境科学, 2014, 35(4): 1343–1350. |
| [${referVo.labelOrder}] | Zheng Z, Liu C l, Lan J, et al. 2014. Factorial analysis of root canal irrigants and cements on coronal microleakage after cementing metal post-cores[J]. West China Journal of Stomatology, 32(3): 242–245. |