 2017, Vol. 37
2017, Vol. 37



2. 上海海洋大学海洋科学学院, 上海 201306;
3. 大洋渔业资源可持续开发省部共建教育部重点实验室, 上海 201306;
4. 江苏省海洋与渔业局, 南京 210003
2. College of Marine Sciences, Shanghai Ocean University, Shanghai 201306;
3. Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Ministry of Education, Shanghai Ocean University, Shanghai 201306;
4. Ocean and Fishery Bureau of Jiangsu, Nanjing 210003
海水中的营养盐是浮游植物的重要营养来源, 营养盐浓度和结构的改变会对浮游植物群落产生直接影响.而浮游植物是海洋中低营养级的主体, 其种群和数量的改变会导致整个海洋生态系统结构和功能的变化.营养盐浓度过低会限制浮游植物的生长与繁殖, 浓度过高或营养盐结构不合理又会导致海水富营养化, 严重危害海洋生态系统的正常运行和功能发挥(Danielsson et al., 2008; Abe et al., 2015).Hecky等(1988)和Dortch等(1982)根据海洋硅藻吸收的营养盐成分比率及海水可利用营养盐的溶解形态, 提出了浮游植物生长中可能出现的营养盐限制条件. Justić等(1995)根据浮游植物生长所需环境溶解营养盐的原子比, 提出了系统评估每一种营养盐化学计量限制的标准.这种方法得到了学者们的广泛应用(张均顺等, 1995; 孙晓霞等, 2011; 宋秀贤等, 2013).
海州湾位于苏鲁交界处, 是江苏近海重要的渔场之一, 但由于过度捕捞、环境污染和鱼虾产卵场等栖息地的破坏, 传统经济鱼类如大黄鱼、小黄鱼等已难以形成渔汛(章守宇等, 2006).海州湾属于开敞型海湾, 发育较宽的潮滩和平缓的海底平原(陈则实, 2007)为海洋牧场建设和人工鱼礁投放提供了有利条件.为了修复和保护海州湾生态环境, 农业部渔业局和江苏省海洋渔业局共同组织实施了"江苏省海洋牧场示范区建设"项目.该项目于2007年启动, 截至2015年已形成了面积为170.25 km2的海洋牧场区域.
针对海洋牧场生态修复的研究, 目前主要集中在生物资源与结构组成调查、沉积物中氮磷等营养盐季节变化与形态分析等方面(张迎秋等, 2016; 张硕等, 2015; 高春梅等, 2015).有关海洋牧场人工鱼礁投放后, 营养盐浓度与结构的变化趋势的研究鲜见报道.本文以2008—2015年共24个航次的调查数据为基础, 通过海州湾海洋牧场鱼礁区和对照区活性磷酸盐(PO43--P)、活性硅酸盐(SiO32--Si)及总溶解无机氮(DIN)的变化对比和营养盐与人工鱼礁投放量的相关性分析, 研究营养盐的浓度与分布特征, 初步探讨海洋牧场营养盐结构及对浮游植物生长的可能限制因素, 尝试较为综合地评估人工鱼礁建设对该海域营养盐的影响, 为人工鱼礁的建设与管理提供科学依据.
2 材料和方法(Materials and methods) 2.1 调查站位在海州湾海域内(34°48'~34°58'N, 119°20'~119°35'E), 共设置9个采样站点(图 1).其中, 海洋牧场区站点6个(RA1~RA6), 对照区站点3个(CA1~CA3), 鱼礁区站点选取在鱼礁投放区, 对照区站点选取在海洋牧场外围.每次采样均采用GPS定位仪对采样站点定位.于2008—2015年, 分别在每年的5月(春季)、8月(夏季)、10月(秋季)进行共计24个航次的调查.
 |
| 图 1 海州湾海洋牧场采样站点示意图 Fig. 1 Sampling stations in marine farming zone of Haizhou Bay |
用有机玻璃采水器采集表层海水, 4 ℃下冷藏保存, 带回实验室分析.营养盐指标NO3--N、NO2--N、NH4+-N、PO43--P、SiO32--Si的实验分析方法严格按照《海洋监测规范》中标准GB173378.3—2007进行.实验数据采用SPSS19.0、Origin8.0等软件进行统计分析与图表制作.
2.3 营养盐结构及限制因素评价本文选用Justić等(1995)建立的化学计量比和营养盐限制因素标准评估该海域的营养盐结构及浮游植物生长的可能限制因素:①Si/P>22和N/P>22, 磷酸盐为限制因素; ②N/P<10和Si/P>1, DIN为限制因素; ③Si/P<10和Si/N<1, 硅酸盐为限制性因素.基于对营养盐吸收动力学研究, 以Si=2 μmol·L-1、DIN=1 μmol·L-1、P=0.1 μmol·L-1作为浮游植物生长的最低阈值(Brown et al., 1979; Nelson et al., 1992).
3 结果与讨论(Results and discussion) 3.1 人工鱼礁投放海州湾海洋牧场的建设以人工鱼礁的投放为主, 2010年的投放规模最大为25647 m3·空, 且投放种类不再单一化, 截至2015年累积投放人工鱼礁达129491 m3·空, 具体投放情况列于表 1中.为改善海洋牧场区原泥沙基质类型, 选择以钢筋混凝土和原石为主材的礁体.鱼礁类型以渔场修复鱼礁为主, 礁体形状包括三角形、方形、十字形和塔形石块礁等.投放方式采用集中堆积, 单座堆积体高度在1.5 m以上.C3D型高分辨侧扫声纳水下扫描结果显示, 已投放的鱼礁底部周围出现了沉积物堆积, 这可能是海州湾海洋牧场处于潮流运动内外的缓冲区, 而人工鱼礁对沉积物输运有一定的滞缓作用(狄欢等, 2013).
| 表 1 2007—2014年海州湾人工鱼礁投放量 Table 1 Delivery volume of artificial fish reefs during 2007—2014 |
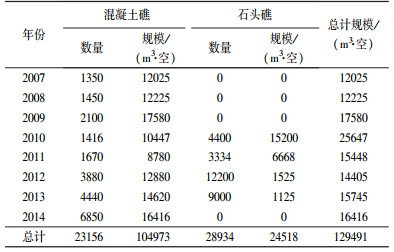
表 2列出了海州湾海洋牧场鱼礁区与对照区春、夏和秋3个季节营养盐浓度的统计结果.从中可以看出, 鱼礁区PO43--P在春、夏、秋季平均浓度分别为0.12、0.15与0.18 μmol·L-1, 对照区PO43--P在春、夏、秋季平均浓度分别为0.09、0.12、0.14 μmol·L-1.鱼礁区SiO32--Si在春、夏、秋季平均浓度分别为9.02、9.12、10.95 μmol·L-1, 对照区SiO32--Si在春、夏、秋季平均浓度分别为8.47、8.69、10.37 μmol·L-1.鱼礁区PO43--P和SiO32--Si浓度高于对照区.海州湾海洋牧场离岸较远, 海水营养盐变化主要受潮流动力和地貌地形控制(赵建华等, 2015).人工鱼礁的投放增加了该海域地质地貌的异质化程度, 形成的流态效应将底层水营养物质带至表层海水(刘同渝, 2003; 林军等, 2006), 造成了鱼礁区营养盐的浓度要高于对照区.
| 表 2 2008—2015年海州湾海洋牧场营养盐浓度统计结果 Table 2 Statistical results of nutrient contents in marine farming zone of Haizhou Bay during 2008—2015 |
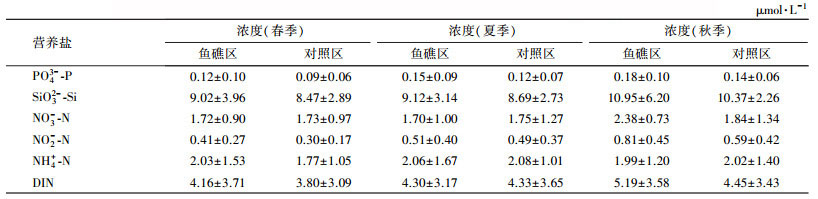
鱼礁区DIN平均浓度在秋季为5.19 μmol·L-1, 较春季(4.16 μmol·L-1)、夏季(4.30 μmol·L-1)增长明显, 对照区DIN浓度也呈现出秋季>夏季>春季的特点.鱼礁区和对照区的PO43--P、SiO32--Si季节变化与DIN相同.海洋牧场示范区海域水深在15 m左右, 秋季海州湾风浪较大, 潮汐和水体扰动作用增强, 人工鱼礁流态效应加剧, 上下层海水交换更频繁, 致使该海域秋季的营养盐浓度要高于春季和夏季.
3.3 人工鱼礁对鱼礁区与对照区营养盐变化的影响海州湾海洋牧场PO43--P和SiO32--Si变化见图 2, 图 3列出了不同季节海洋牧场DIN变化.在海洋牧场建设期间, 鱼礁区和对照区的营养盐出现了不同程度的变化.鱼礁区PO43--P浓度呈增长趋势, 尤以夏、秋两季较为明显(图 2a).对照区秋季PO43--P稳定且高于春季和夏季, 春季和夏季的年际波动范围较大.鱼礁区和对照区SiO32--Si的年际变化基本一致(图 2b), 2010年后SiO32--Si浓度都呈现出秋季大于春、夏两季的趋势.鱼礁区和对照区春、夏和秋季DIN浓度呈现整体增长趋势(图 3), DIN中不同形态的组成也发生了改变.2012年之前, NO3--N为DIN的主要组分, 所占比例可达40%~70%.2012年后, NO3--N所占比例下降, NH4+-N所占比例由原来的10%~50%上升至25%~60%.NH4+-N浓度的增加使其成为DIN的主要组分.NO2--N占DIN总量的15%左右, 在整个过程中变化不大.鱼礁区和对照区DIN组分形态变化相一致, 但在浓度上鱼礁区要高于对照区, 2012年后对照区DIN浓度出现明显增长.
 |
| 图 2 2008—2015海州湾海洋牧场春季、夏季和秋季鱼礁区与对照区PO43--P(a)和SiO32--Si(b)的年份变化 Fig. 2 The changes of PO43--P(a) and SiO32--Si(b) in reef and control areas of Haizhou Bay during spring, summer and autumn in 2008—2015 |
 |
| 图 3 2008—2015海州湾海洋牧场春季(a)、夏季(b)和秋季(c)鱼礁区与对照区DIN变化 Fig. 3 The changes of DIN in reef and control areas of Haizhou Bay during spring(a), summer(b) and autumn(c) in 2008—2015 |
沉积物营养盐的释放是海洋牧场水体营养盐的重要来源(高春梅等, 2016).近年来海洋牧场区域筏式吊养藻类的养殖面积不断增加, 藻类生长死亡和其他生物活动所产生的生物碎屑可能会导致海洋牧场海水和沉积物中的营养盐浓度的增加(陈菊芳等, 2006).人工鱼礁所形成的流场效应在增加上下层海水交换的同时, 还能够将沉积物携带至底层海水(王伟定等, 2010; Yanagi et al., 1991), 沉积物营养盐向海水中的释放能持续提高海水中营养盐的浓度.
为了探讨鱼礁累积投放对营养盐的影响, 分析了鱼礁投放量与营养盐浓度的相关性, 结果列于表 3.从表 3可知, 鱼礁区PO43--P与人工鱼礁累积投放量呈显著正相关(r=0.861, p < 0.01), 表明鱼礁区PO43--P在很大程度上可能受到了鱼礁投放量的影响.对照区PO43--P与鱼礁累积投放量的相关性不显著(r=0.118), 说明人工鱼礁投放对对照区的影响较鱼礁区要小.韩照祥等(2012)对海州湾及其邻近海域沉积物-水界面的磷形态特征与动力学研究表明, 生物扰动和微生物的分解作用会加剧PO43--P在沉积物-水界面上的交换作用.对海洋牧场鱼礁区底栖生物的调查表明, 人工鱼礁的投放增加了底栖生物的生物量与种类(张虎等, 2008), 加上人工鱼礁投放造成生物扰动的加剧可能是鱼礁区PO43--P浓度增加的主要原因之一.海州湾为半日潮, 自身潮流比较弱, 不利于营养盐的运送(周德山等, 2008), 导致人工鱼礁建设对对照区的影响较小.
海水中的SiO32--Si主要来源于陆地径流的输入(Harvey, 1956).海洋牧场区离海岸较远, 从临洪河、青口河与龙王河等入海河口输入的SiO32--Si到达海洋牧场鱼礁区和对照区时, 浓度差异已不显著.从表 3看出, SiO32--Si与鱼礁累积投放量的相关性不显著(鱼礁区r=0.179, 对照区r=-0.084), 说明海洋牧场底部的人工鱼礁对陆源输入的SiO32--Si影响较小.因此, 鱼礁区与对照区SiO32--Si的年际变化趋势基本一致.2010年后海洋牧场SiO32--Si浓度呈现出秋季大于春、夏两季的趋势.海洋牧场浮游植物的调查结果表明, 浮游植物以硅藻门为主, 秋季浮游植物种类和丰度均远高于春、夏两季(杨晓改等, 2014).秋季SiO32--Si的增加促进了该海域浮游植物的生长, 这佐证了海洋牧场SiO32--Si的变化情况.
| 表 3 海洋牧场营养盐与鱼礁累积投放量的相关性 Table 3 Correlation coefficient between nutrients and accumulated amount of fish reef in marine farming zone |
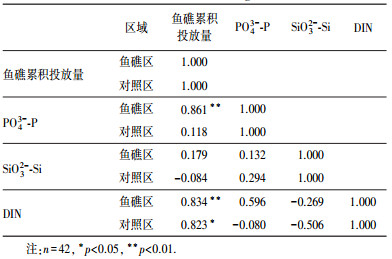
海洋牧场鱼礁区和对照区DIN随年际变化呈整体增长趋势.鱼礁区和对照区DIN与鱼礁累积投放量都呈显著正相关, 对照区DIN与鱼礁累积投放量呈显著正相关(r=0.823, p < 0.05).不同于SiO32--Si, 海洋牧场鱼礁区DIN的年际变化与PO43--P类似, 在很大程度上受到了鱼礁投放的影响.沉积物可以为海洋牧场提供124%的DIN营养供给(高春梅等, 2016).人工鱼礁的流场效应和生物扰动, 会加剧沉积物中的氮营养盐向下层海水释放(刘同渝等, 2003).上下层水体的交换, 增加了鱼礁区表层海水中的DIN浓度.当氨化作用和硝化作用进行充分时, NO3--N、NH4+-N和NO2--N会达到基本的热力学平衡.鱼礁区DIN浓度升高时, 不同形态氮之间的热力学过程会造成对照区DIN浓度的增加, 因而随着人工鱼礁的建设, 对照区DIN浓度呈明显增长趋势.同时, 鱼礁区和对照区在2012年后NH4+-N取代NO3--N成为DIN的主要组分.人工鱼礁的逐年投放, 使沉积物中的氮成为海洋牧场的稳定源.沉积物向水体中释放的氮是随鱼礁投放不断增加的, 而浮游植物会优先吸收水体中的NO3--N(韩丽君等, 2015).海洋牧场藻类养殖区的扩大加快了NO3--N的消耗.因此, 导致海洋牧场DIN组分发生变化, 表现为NH4+-N浓度的增加.
3.4 海洋牧场鱼礁区与对照区营养盐结构变化图 4给出了2008—2015年间海州湾海洋牧场营养盐结构的变化情况.从图 4可以看出, 鱼礁区和对照区Si/P、DIN/P、Si/DIN的年际变化趋势基本一致.但随着时间变化, Si/P与DIN/P的比值由鱼礁区>对照区变为鱼礁区 < 对照区.人工鱼礁的建设提升了鱼礁区的PO43--P浓度, SiO32--Si在鱼礁区和对照区的变化一致, 对照区SiO32--Si和PO43--P受人工鱼礁建设的影响小.因此, Si/P、DIN/P由鱼礁区>对照区转变为鱼礁区 < 对照区.Si/DIN呈波动状态, 自2010年后Si/DIN有趋于稳定的趋势, 且接近于Redfield比值1.总体而言, 海州湾海洋牧场示范区的Si/P、DIN/P处于较高比值, 与Redfield平衡状态相差较大, Si/DIN比值不高(0.2~5.0), 这与谢琳萍等(2012)的调查结果基本一致.
 |
| 图 4 2008—2015年间海州湾海洋牧场营养盐结构变化 Fig. 4 The changes of nutrient ratios in the marine farming zone of Haizhou Bay during 2008—2015 |
利用Justić法评估海洋牧场的营养盐结构及浮游植物生长的可能限制因素时发现(表 4), 海洋牧场鱼礁区PO43--P作为限制因子出现的概率最高(春季56.8%, 夏季47.7%, 秋季66.7%), 由于浓度过低原因造成的限制出现的概率高(春季27.3%, 夏季36.4%, 秋季31.0%).对照区限制因子出现概率最高的同样为PO43--P(春季50.0%, 夏季45.8%, 秋季62.5%), 由于浓度的限制所占比例同样很高(春季30.0%, 夏季30.0%, 秋季41.7%).鱼礁区DIN作为限制因子出现时(春季11.4%, 夏季18.2%, 秋季9.5%), DIN浓度起到了一定的限制作用(所占比例为春季4.5%, 夏季9.1%, 秋季2.1%).对照区DIN作为限制因子出现时(春季8.3%, 夏季29.2%, 秋季4.2%), 春季是主要受DIN浓度的影响(所占比例8.3%), 夏季和秋季不受DIN浓度的影响(夏季0, 秋季0), 受控于营养盐结构.SiO32--Si在鱼礁区和对照区的春季均不作为限制因子出现, 夏季作为限制因子出现时不受浓度的影响(鱼礁区11.4%, 对照区8.3%), 秋季作为限制因子出现时受浓度控制(鱼礁区9.5%, 对照区12.5%).
| 表 4 海州湾海洋牧场各种营养盐限制因子出现的概率 Table 4 The occurrence probabilityof nitrogen, phosphorus and silicon in marine farming of Haizhou Bay during different seasons |
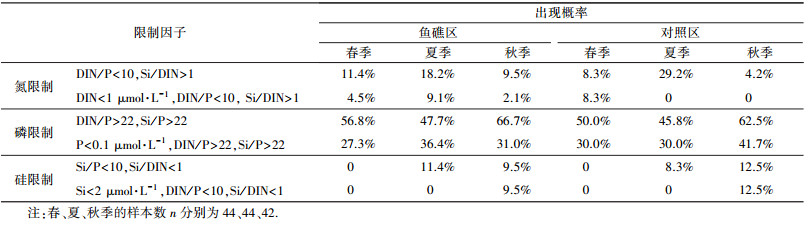
海州湾海洋牧场营养盐对浮游植物多表现为磷限制特征, 赵建华等(2105) 和倪金俤等(2011)在研究海州湾营养盐结构时, 也得出了海州湾整体表现为明显的磷限制的结论.海洋牧场人工鱼礁的投放虽然增加了该海域水体中的PO43--P浓度, 但鱼礁区和对照区由于PO43--P浓度所造成的营养盐限制比例仍然很大.这表明海洋牧场的营养盐结构不仅受人工鱼礁投放的影响, 还受到整个海湾环境的影响.当DIN作为限制因子出现时, DIN浓度对鱼礁区的影响要大于对照区, 尤其夏季和秋季更为明显.对海洋牧场区域浮游植物的调查结果显示, 鱼礁区和对照区浮游植物在群落结构和丰度上无显著差异(刘长东等, 2015), 说明鱼礁区和对照区DIN均能满足浮游植物的生长要求.DIN作为限制因子的出现, 可能是由于在海洋牧场人工养殖的藻类消耗水体中营养盐造成的.SiO32--Si对鱼礁区和对照区的限制作用相同, 差异性主要体现在季节变化上.海洋牧场中的浮游植物以占84.3%的硅藻门为主, 硅藻生长需要消耗海水中的SiO32--Si, 而SiO32--Si主要来自陆源输入, 受人工鱼礁建设的影响不大.硅藻的季节性生长可能控制着海洋牧场SiO32--Si的变化, 因此导致不同季节SiO32--Si的限制作用发生变化.
4 结论(Conclusions)1) 海洋牧场鱼礁区和对照区PO43--P、SiO32--Si浓度呈现出秋季>夏季>春季, 且鱼礁区浓度高于对照区.鱼礁区DIN浓度在秋季最高, 且比春、夏季增长显著; 对照区DIN浓度也呈现出秋季>夏季>春季, 但鱼礁区DIN浓度也高于对照区.
2) 人工鱼礁的投放量增加了鱼礁区PO43--P和DIN浓度, 受热力学平衡的影响, 对照区DIN浓度也增加, 对照区PO43--P浓度受人工鱼礁投放的影响较小, 人工鱼礁投放对SiO32--Si影响不大.
3) 鱼礁区和对照区DIN主要组成发生改变, 自2012年, 主要组成由NO3--N变为NH4+-N.
4) 海洋牧场鱼礁区和对照区对浮游植物生长的限制主要表现为PO43--P限制; DIN成为限制因子时, 对鱼礁区的影响大于对照区, 尤其是在夏、秋季节; SiO32--Si对鱼礁区和对照区限制作用类似.
Abe K, Tsujino M, Nakagawa N, et al. 2015. Characteristic of Si:P:N ratio in bottom water in central Suo-Nada, western Seto Inland Sea[J]. Journal of Oceanography, 71(1): 53–63.
DOI:10.1007/s10872-014-0262-4
|
Brown E J, Button D K. 1979. Phosphate-limited growth kinetics of Selenastrumcapricornutum(Chlorophyceae)[J]. Journal of Phycology, 15(3): 305–311.
DOI:10.1111/jpy.1979.15.issue-3
|
陈菊芳, 齐雨藻, 徐宁, 等. 2006. 大亚湾澳头水域浮游植物群落结构及周年数量动态[J]. 水生生物学报, 2006, 30(3): 311–317.
|
陈则实. 2007. 中国海湾引论[M]. 北京: 海洋出版社: 337–338.
|
Danielsson A, Papush L, Rahm L. 2008. Alterations in nutrient limitations-scenarios of a changing Baltic Sea[J]. Journal of Marine Systems, 73(3/4): 263–283.
|
Dortch Q. 1982. Effect of growth conditions on accumulation of internal nitrate, ammonium and protein in three marine diatoms[J]. Journal of Experimental Marine Biology & Ecology, 61(3): 243–264.
|
狄欢, 张硕, 钱卫国. 2013. 海州湾海洋牧场区表层沉积物主要理化状况及其相关性分析[J]. 大连海洋大学学报, 2013, 28(4): 406–412.
|
高春梅, 郑伊汝, 张硕. 2016. 海州湾海洋牧场沉积物-水界面营养盐交换通量的研究[J]. 大连海洋大学学报, 2016, 31(1): 95–102.
|
高春梅, 朱珠, 王功芹, 等. 2015. 海州湾海洋牧场海域表层沉积物磷的形态与环境意义[J]. 中国环境科学, 2015, 35(11): 3437–3444.
DOI:10.3969/j.issn.1000-6923.2015.11.031 |
Harvey H W. 1956. The chemistry and fertility of sea waters[J]. The Quarterly Review of Biology, 31(3): 176–176.
|
Hecky R E, Kilham P. 1988. Nutrient limitation of phytoplankton in freshwater and marine environments: A review of recent evidence on the effects of enrichment[J]. Limnology and Oceanography, 33(4part2): 796–822.
DOI:10.4319/lo.1988.33.4part2.0796
|
Justić D, Rabalais N N, Turner R E, et al. 1995. Changes in nutrient structure of river-dominated coastal waters: stoichiometric nutrient balance and its consequences[J]. Estuarine Coastal & Shelf Science, 40(3): 339–356.
|
韩丽君, 郑新庆, 蓝文陆, 等. 2015. 近10年涠洲岛周边海域表层海水营养盐浓度变化特征[J]. 应用海洋学学报, 2015, 34(1): 65–72.
|
韩照祥, 王超, 朱圳, 等. 2012. 海州湾及其临近海域沉积物-水界面的磷形态特征与动力学[J]. 环境科学与技术, 2012(11): 12–15.
DOI:10.3969/j.issn.1003-6504.2012.11.003 |
林军, 章守宇. 2006. 人工鱼礁物理稳定性及其生态效应的研究进展[J]. 海洋渔业, 2006, 28(3): 257–262.
|
刘同渝. 2003. 人工鱼礁的流态效应[J]. 江西水产科技, 2003(6): 43–44.
|
刘永虎, 程前, 田涛, 等. 2016. 大连獐子岛人工鱼礁海域夏季水质变化与评价[J]. 大连海洋大学学报, 2016, 31(3): 331–337.
|
刘长东, 郭晓峰, 唐衍力, 等. 2015. 海州湾前三岛人工鱼礁区浮游植物群落组成及与环境因子的关系[J]. 中国水产科学, 2015, 22(3): 545–555.
|
Nelson D M, Treguer P. 1992. Role of silicon as a limiting nutrient to Antarctic diatoms: Evidence from kinetic studies in the Ross Sea ice-edge zone[J]. Marine Ecology Progress, 80(2/3): 255–264.
|
倪金俤, 矫新明, 盖建军, 等. 2011. 营养元素对海州湾藻类生长的影响[J]. 水产养殖, 2011, 32(1): 34–37.
|
宋秀贤, 俞志明, 殷克东, 等. 2013. 香港近岸海域营养盐结构特征及其对浮游植物生长的影响[J]. 海洋与湖沼, 2013, 44(4): 846–852.
|
孙晓霞, 孙松, 赵增霞, 等. 2011. 胶州湾营养盐浓度与结构的长期变化[J]. 海洋与湖沼, 2011, 42(5): 662–669.
DOI:10.11693/hyhz201105006006 |
王伟定, 梁君, 章守宇. 2010. 人工鱼礁建设对浙江嵊泗海域营养盐与水质的影响[J]. 水生生物学报, 2010, 34(1): 78–87.
|
谢琳萍, 孙霞, 王保栋, 等. 2012. 渤黄海营养盐结构及其潜在限制作用的时空分布[J]. 海洋科学, 2012, 36(9): 45–53.
|
Yanagi T, Nakajima M. 1990. Change of oceanic condition by the man-made structure for upwelling[J]. Marine Pollution Bulletin, 23(4): 131–135.
|
杨晓改, 薛莹, 昝肖肖, 等. 2014. 海州湾及其邻近海域浮游植物群落结构及其与环境因子的关系[J]. 应用生态学报, 2014, 25(7): 2123–2131.
|
张虎, 刘培廷, 汤建华, 等. 2008. 海州湾人工鱼礁大型底栖生物调查[J]. 海洋渔业, 2008, 30(2): 97–104.
|
张均顺, 沈志良. 1997. 胶州湾营养盐结构变化的研究[J]. 海洋与湖沼, 1997, 28(5): 529–535.
|
张硕, 王功芹, 朱珠, 等. 2015. 海州湾表层沉积物中不同形态氮季节性赋存特征[J]. 生态环境学报, 2015, 24(8): 1336–1341.
|
张迎秋, 许强, 徐勤增, 等. 2016. 海州湾前三岛海域底层鱼类群落结构特征[J]. 中国水产科学, 2016, 23(1): 156–168.
|
章守宇, 张焕君, 焦俊鹏, 等. 2006. 海州湾人工鱼礁海域生态环境的变化[J]. 水产学报, 2006, 30(4): 475–480.
|
赵建华, 李飞. 2015. 海州湾营养盐空间分布特征及影响因素分析[J]. 环境科学与技术, 2015, 38(12): 32–35.
|
周德山. 2008. 海州湾海域赤潮形成的环境因子研究[D]. 苏州: 苏州大学. 1-8
http://cdmd.cnki.com.cn/article/cdmd-10285-2008127188.htm |