 2017, Vol. 37
2017, Vol. 37



2. 中国科学院水利部水土保持研究所, 杨凌 712100
2. Institute of Soil and Water Conservation, Chinese Academy of Sciences and Ministry of Water Resources, Yangling 712100
土壤呼吸是陆地生态系统碳循环的重要组成部分.研究表明, 以土壤呼吸为主体的CO2排放的增加可能会加剧气候变化, 并已成为全球气候变化研究中的核心内容之一(Schlesinger et al., 2000).土壤呼吸是反映土壤活性和生态系统生产力的重要指标, 主要包括根系自养呼吸和土壤微生物异养呼吸, 其大小既受土壤温度、土壤水分、降雨和土壤理化性质等非生物因子的影响, 也受植被类型、物种属性、根系生物量等生物因子的影响, 具有明显的时空变异性特征(Tang et al., 2005; Xu et al., 2001a).
在半干旱地区, 降雨是土壤水分的主要来源, 降雨事件的发生与否会改变土壤的水分和通气状况, 影响土壤呼吸底物的组成与有效性, 因此, 土壤呼吸对水分的变化极为敏感(Raich et al., 2000; Gestel et al., 1993).研究表明, 全球和区域内的水分循环和降雨格局正在改变(Ball et al., 1999), 如我国北方地区的降水有增加的趋势(Zhang et al., 2006), 且大降雨事件趋于频繁(Liu et al., 2005a).降雨格局的改变会影响土壤水分的变动模式(如土壤干旱与湿润的时间与程度, 以及干湿交替的频度等)(Jackson et al., 2001), 进而影响植被生长和植被群落构成(Knapp et al., 2001), 对土壤呼吸的各组分产生影响.其中, 植被对土壤呼吸的影响主要包括拦截太阳辐射从而影响土壤能量平衡, 截留降雨和蒸腾吸水调节土壤水分等间接效应(Rutter et al., 1977), 以及植被枯落物为微生物呼吸提供底物(Gillon et al., 1999)和光合作用影响根系呼吸强度等直接效应(Huxman et al., 2004).在较小的时间和空间尺度上, 降雨量对土壤呼吸的影响因素较为复杂(彭琴等, 2012), 如小降雨事件能够迅速增加表层土壤的异养呼吸, 该激发效应随水分的蒸发而减弱(Knapp et al., 2008); 大降雨发生后, 水分入渗到更深层土壤, 会激发植物根系的自养呼吸(Ryana, 2007).目前, 关于土壤呼吸及其组分对模拟降雨的响应已有诸多研究(Sponseller, 2007; Cable et al., 2008; Qi et al., 2014), 但关于降雨量对土壤呼吸速率的影响与群落类型的关系研究尚少.深入研究不同类型植被下土壤呼吸对降雨的响应过程与差异, 将有助于揭示降雨对植被生态系统碳交换过程的影响与机制(Travis et al., 2004).
黄土丘陵区是典型生态脆弱区, 也是我国水土流失严重的区域之一(Fu et al., 2000).自实行退耕还林还草工程以来, 大范围的坡耕地转变成天然草地.典型草地优势种在群落中相对比例的高低不仅反映了退耕草地的演替阶段, 也往往决定了其CO2的排放特征(温仲明等, 2005).目前, 对黄土丘陵区天然草地的研究多集中在群落分布、物种生物学属性、土壤性状特征等方面(张娜等, 2004; 步秀芹等, 2007; 牛富荣等, 2011), 而对灌丛草地碳交换机理及其与环境要素的关系等报道较少(黄懿梅等, 2009).因此, 本研究选取黄土丘陵区3种典型天然草地群落, 分别代表不同退耕年限的草地类型, 通过模拟降雨试验, 比较黄土丘陵区天然退耕草地群落土壤呼吸对次降雨的响应规律及其动态变化特征, 阐明不同优势种草地群落土壤呼吸对不同等级降雨量的短期响应过程与差异, 以期为全面评估黄土丘陵半干旱区退耕草地生态系统碳收支提供基础参数.
2 材料与方法(Materials and methods) 2.1 研究区概况试验地位于陕西安塞农田生态系统国家野外科学观测研究站, 地理位置为东经109°19′23″, 北纬36°51′31″, 海拔为1068~1309 m.该区年平均温度为8.8 ℃, 最冷月1月平均温度为-6.9 ℃, 最热月7月平均温度为22.6 ℃, 全年≥10 ℃积温为3113.9 ℃, 无霜期159 d, 多年平均降雨量510 mm, 降雨季节分布不均, 7—9月降雨量占全年降水量的60%~80%.其中2005—2013年间, 6月降雨量占全年降水量的9.1%, 7月降雨量占全年降水量的29.6%.气候属暖温带半干旱气候, 植被属暖温带半干旱森林草原区, 处于暖温带落叶阔叶林区向暖温带草原区的过渡带.土壤以黄绵土、绵沙土等为主.地带性植被为森林灌丛草原, 具有明显的过渡性特色; 天然草地主要为灌木草丛、干草原及低湿草甸等; 典型地带性草本植物有白羊草(Bothriochloa ischaemum (Linn.) Keng)、达乌里胡枝子(Lespedeza davurica (Laxm.) Schindl)、铁杆蒿(Artemisia gmelinii Web.et Stechm.)、长芒草(Stipa bungeana Trin.)、茭蒿(Artemisia giraldii Pamp.)等.其中, 白羊草为禾本科孔颖草属多年生草本C4植物, 广泛分布在黄土丘陵区的低山丘陵、梁峁顶部的暖温带地段, 是显域性植被建群种; 达乌里胡枝子是豆科达乌里胡枝子属多年生草本状半灌木, 为优良的乡土牧草之一, 是白羊草灌丛草地的亚优势种; 铁杆蒿为暖温性中旱生半灌木植物, 在退耕后逐渐成为群落优势种并稳定存在(唐龙等, 2006).
2.2 研究方法 2.2.1 小区设计试验小区设置在山地试验场坡地, 坡度为24°, 坡向为南偏东10°.天然草地群落以白羊草、达乌里胡枝子和铁杆蒿为优势种, 分别选取以上3种植物为主要优势种的1 m×1 m的样方各15个.
白羊草群落样地位于坡地上半部, 6月植被盖度为59.34%, 白羊草占20.32%, 7月植被盖度为70.04%, 白羊草占29.44%;达乌里胡枝子群落样地位于下坡段西边, 白羊草和达乌里胡枝子均匀分布, 同为群落优势种, 该样地6月植被盖度为54.12%, 达乌里胡枝子占9.45%, 7月植被盖度为73.16%, 达乌里胡枝子占11.31%.铁杆蒿群落样地位于下坡段东边, 铁杆蒿生长旺盛, 为主要优势种, 该区6月植被盖度为57.40%, 铁杆蒿占23.45%, 7月植被盖度为75.32%, 铁杆蒿占30.17%.
2.2.2 降雨处理按照陕西黄土高原地区日降雨量, 降雨事件可划分为小雨事件(0.1~9.9 mm)、中雨事件(10~24.9 mm)、大雨事件(25~49.9 mm)和暴雨事件(≥50 mm) (孙智辉等, 2009).通过统计2005—2013年期间降雨资料, 安塞站6月和7月不同等级降雨量频次分别为:0~5 mm降雨发生频次最高, 但难以形成有效穿透雨; 5 mm以上降雨事件多为10~20 mm的中雨, 且月份分布均匀; 50 mm以上极端降雨有增多趋势.因此, 依照该降雨频次与分级, 本研究设置15 mm降雨以模拟发生频次较高的中雨事件, 并设置50 mm降雨以模拟暴雨(极端降雨)事件, 具体将降雨量分别为0(未降雨、对照)、15和50 mm.每个处理重复3次.
降雨试验分别于2014年6月16日和7月16日进行, 为减少蒸发损失和风的影响, 模拟降雨开始于当日19:00.采用便携式野外人工模拟降雨器(专利号:2013103763306), 降雨器由压力设备、储水槽、碎流挡板、喷头和支撑设备组成.根据试验操作的实际情况和可行性, 设置平均降雨强度为0.5 mm·min-1, 降雨器喷头高度约2.5 m.
2.2.3 测定项目与方法模拟降雨当天及降雨后连续7 d对土壤温度、土壤水分和土壤呼吸速率进行监测, 测定于每天上午9:00—11:00进行, 该时间段土壤呼吸速率能够代表全天的平均值(Xu et al., 2001b).
土壤温度:采用直角地温计测定, 与土壤呼吸速率测定同步, 为5 cm土层温度; 于基座周围3 cm处随机选取3点测量, 取平均值.
土壤水分:采用Spectrum TDR100便携式水分测定仪测定, 深度为10 cm, 结果以体积含水量表示.
土壤呼吸速率:采用PP System公司产EGM-4便携式CO2分析仪测定, 各小区分别测定, 随机取样, 3次重复; 测定时在尽量不扰动土壤和枯落物的前提下, 将基座嵌入土壤3 cm, 基座为直径10 cm、高度15 cm的不锈钢圆形筒.
2.3 数据处理采用SPSS 20.0软件对数据进行统计分析, 运用多因素方差分析(Three-way ANOVA)检验月份、降雨量和群落类型对土壤温度和水分的影响, 用单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)检验各降雨量处理间土壤呼吸差异(显著性水平α=0.05).运用SigmaPlot 12.5软件绘图.
土壤温度与土壤呼吸速率间的关系采用指数模型Rs=a·ebTs进行拟合, 其中, Rs为土壤呼吸速率(g·m-2·h-1), Ts为5 cm土层土壤温度(℃).土壤呼吸温度敏感性系数Q10值采用指数关系模型Q10=e10b计算.
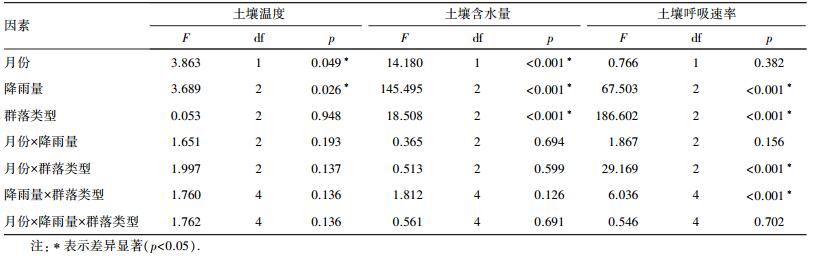
3 结果与分析(Results and analysis) 3.1 土壤温度和土壤含水量土壤温度在不同月份和不同降雨处理间差异显著(表 1).6月, 不同降雨处理下3种草地群落土壤温度均呈先降低后上升趋势, 变化范围是20.5~32.6 ℃(图 1).模拟降雨后1~3 d, 白羊草和达乌里胡枝子群落平均土壤温度较对照处理分别降低了9.8%和15.8%, 铁杆蒿群落内降雨处理下土壤温度与对照无显著差异.降雨后第4 d起, 白羊草群落土壤温度在各降雨处理组间均无显著差异, 达乌里胡枝子群落15 mm降雨处理和对照组土壤温度无显著差异, 并且均显著高于50 mm处理.7月, 3种草地群落土壤温度波动范围是19.0~34.3 ℃, 均呈逐渐下降趋势.白羊草和达乌里胡枝子群落中, 50 mm降雨处理组土壤温度在降雨后1~3 d显著低于对照, 第4 d起降雨处理组土壤温度与对照无显著差异.不论是6月还是7月, 铁杆蒿群落土壤温度在各降雨处理组间无显著差异(p>0.05).
| 表 1 月份、降雨量和群落类型对土壤温度、土壤含水量和土壤呼吸速率的影响(n=3) Table 1 Analysis of variance for the effects of growth month, rainfall size, community type on soil temperature, soil water content and soil respiration rate |
 |
| 图 1 模拟降雨前后3种草地群落土壤温度动态变化(横坐标中0表示模拟降雨前1天; 折线内与折线下方误差线分别为标准误与LSD值(α=0.05, n=3), 下同) Fig. 1 Dynamics of soil temperature in three communities before and after simulated rainfall in June and July, 2014 |
降雨处理间及群落类型间土壤含水量差异显著(表 1).6月, 模拟降雨后3种群落土壤含水量均呈上升趋势(图 2).白羊草群落土壤含水量峰值出现在50 mm降雨处理下的第1 d(8.21%), 达乌里胡枝子和铁杆蒿群落的土壤含水量最高值出现在50 mm降雨后第2 d, 分别为7.40%和7.56%.模拟降雨前, 3种群落土壤含水量分别为:白羊草6.39%, 达乌里胡枝子6.17%, 铁杆蒿5.70%.降雨后7 d内, 3种群落平均土壤含水量均表现为50 mm降雨处理显著高于15 mm降雨处理, 且均显著高于对照组.7月, 3种群落土壤含水量在降雨后第1 d均显著上升, 此后逐渐降低.各群落土壤含水量峰值均出现在降雨后第1 d的50 mm降雨处理组, 分别为:白羊草8.64%, 达乌里胡枝子8.39%, 铁杆蒿8.61%.测定8 d内, 白羊草群落的平均土壤含水量最高(6.43%), 达乌里胡枝子群落和铁杆蒿群落分别为6.16%和5.95%.
 |
| 图 2 模拟降雨前后3种草地群落土壤含水量动态变化 Fig. 2 Dynamics of soil water content in three communities before and after simulated rainfall in June and July, 2014 |
土壤呼吸速率在不同降雨量和群落类型间差异显著, 月份间无差异.月份和群落类型及降雨量和群落类型间的交互作用显著(p < 0.05)(表 1).
6月, 3种群落土壤呼吸速率在降雨后第1 d均显著增加, 此后逐渐下降(图 3).降雨后第1 d, 50 mm降雨处理下白羊草、达乌里胡枝子、铁杆蒿群落土壤呼吸速率分别比降雨前增加了104%、112%和45%;15 mm降雨处理下分别增加了44%、101%和3%.测定期间内, 白羊草群落土壤呼吸速率为0.81~1.65 g·m-2·h-1, 50 mm降雨处理下平均土壤呼吸速率为1.30 g·m-2·h-1, 高于15 mm降雨处理, 且显著高于对照.达乌里胡枝子群落土壤呼吸速率的变化范围是0.52~1.12 g·m-2·h-1, 雨后第1 d, 15和50 mm降雨处理分别升至1.04和1.07 g·m-2·h-1, 测定8 d内, 不同降雨处理下平均土壤呼吸速率无显著差异(p>0.05).铁杆蒿群落土壤呼吸速率变幅为0.66~1.69 g·m-2·h-1, 50 mm降雨后第1 d达到峰值1.70 g·m-2·h-1.测定8 d内, 50 mm处理组平均土壤呼吸速率为1.22 g·m-2·h-1, 显著高于15 mm降雨处理(0.97 g·m-2·h-1)和对照组(0.91 g·m-2·h-1).
 |
| 图 3 模拟降雨前后3种草地群落土壤呼吸速率动态变化 Fig. 3 Dynamics of soil respiration rate in three communities before and after simulated rainfall in June and July, 2014 |
7月, 降雨后第1 d, 50 mm降雨处理下白羊草、达乌里胡枝子、铁杆蒿群落土壤呼吸速率分别比降雨前增加了51%、15%和37%;15 mm降雨处理下分别增加了28%、19%和5%.测定期间, 白羊草群落土壤呼吸速率变化范围是1.01~1.75 g·m-2·h-1, 峰值出现在50 mm降雨后第1 d.模拟降雨后7 d内, 白羊草群落平均土壤呼吸速率为1.25 g·m-2·h-1, 较6月同期增长了15.1%.达乌里胡枝子群落土壤呼吸速率变幅较小, 为0.30~0.99 g·m-2·h-1.降雨后7 d内, 15及50 mm降雨处理组平均土壤呼吸速率与对照组均无显著差异(p>0.05).铁杆蒿群落土壤呼吸速率的波动范围是0.47~1.55 g·m-2·h-1, 均值为0.91 g·m-2·h-1.降雨后第1 d, 50 mm降雨处理的土壤呼吸速率达到峰值1.55 g·m-2·h-1; 15 mm降雨处理的土壤呼吸速率从1.07 g·m-2·h-1升至1.14 g·m-2·h-1; 对照组土壤呼吸速率逐渐降低, 其均值显著低于50 mm降雨处理.
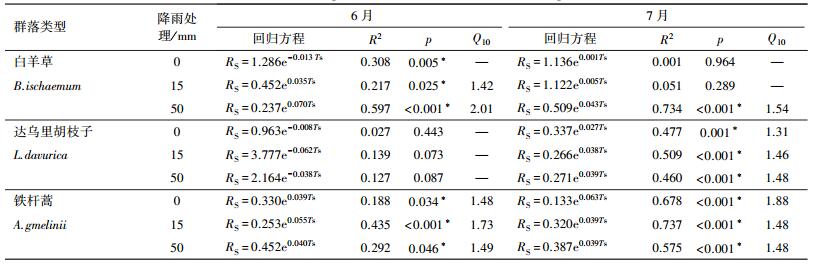
3.3 土壤呼吸温度敏感性系数6月, 达乌里胡枝子群落在各降雨处理下土壤呼吸速率与土壤温度的指数关系均不显著(表 2).根据各降雨处理下土壤呼吸速率与土壤温度指数关系式得知, 白羊草群落15 mm和50 mm降雨处理下Q10值分别为1.42和2.01, 铁杆蒿群落在0、15和50 mm降雨处理下的Q10值分别为1.48、1.73和1.49.7月, 达乌里胡枝子和铁杆蒿群落土壤温度与土壤呼吸速率存在显著的指数关系(p < 0.01), 土壤温度能够解释土壤呼吸变化的46%~74%, 其中, 达乌里胡枝子群落Q10值分别为1.31、1.46和1.48, 铁杆蒿群落Q10值分别为1.88、1.48、1.48.白羊草群落中仅50 mm降雨处理下存在显著指数关系, 对应Q10值为1.54.
| 表 2 各草地群落不同模拟降雨量下土壤呼吸速率与土壤温度的拟合关系 Table 2 Fitting relationships between soil respiration rate (Rs) and soil temperature at 5 cm profile (Ts) in each community under simulated rainfall |
在试验开展的2个月份, 3种草地群落土壤呼吸速率在降雨后第1 d均出现不同程度的提升.其中, 白羊草群落雨后土壤呼吸速率显著提高, 且降雨量越大, 增加量越高; 达乌里胡枝子群落土壤呼吸速率在15 mm和50 mm降雨后均出现小幅提升, 增加量在2种降雨处理间无显著差异; 铁杆蒿群落土壤呼吸速率仅在50 mm降雨后显著提升, 在15 mm降雨后无明显上升, 表明降雨对土壤呼吸有激发效应, 但在不同类型群落中引起土壤呼吸激发的降雨量不同, 呼吸速率增加量也不同.Chen等(2008)在内蒙古羊草草原的模拟降雨试验表明, 土壤呼吸速率在降雨后1 d内达到最大, 且随降雨量的增加, 激发作用增强, 与本文研究结果类似.在干旱半干旱地区, 降雨事件对土壤呼吸的激发效应主要归纳为以下原因:① 降雨后水分入渗到土壤孔隙中置换出长期积聚的CO2, 该过程通常发生在降雨后几分钟至数小时以内(Chen et al., 2008); ② 土壤水分的增加促进了土壤有机质的矿化和地表枯落物的分解, 提高了呼吸底物的有效性(Gestel et al., 1993); ③ 土壤微生物在干旱胁迫下活性较低, 降雨后水分得到补给, 微生物迅速利用底物进行新陈代谢(Grant et al., 1994).本研究中, 为保证土壤水分充分入渗, 土壤呼吸测定在模拟降雨后约12 h进行, 由此可排除物理置换的CO2释放量, 土壤呼吸的激发效应主要与呼吸底物有效性及微生物活性密切相关(Sponseller, 2007).相同降雨条件下, 白羊草群落土壤呼吸速率的增加量及测定期间平均值均显著高于达乌里胡枝子和铁杆蒿群落.研究显示, 黄土丘陵区白羊草群落0~20 cm土层土壤有机碳储量、微生物量、酶活性和多项养分指标显著高于达乌里胡枝子群落, 根系集中分布在0~20 cm土层且须根发达, 为土壤呼吸提供了充足的底物供应(王会梅等, 2005; 李国辉, 2010; 张海东等, 2015; 王百群等, 2010).
3种草地群落土壤呼吸速率与土壤含水量均呈对数函数关系, 白羊草和铁杆蒿群落土壤呼吸速率随土壤含水量增加迅速增加, 达乌里胡枝子提升较缓(图 4).相关研究表明, 土壤含水量增加至接近田间持水量前, 土壤呼吸随水分增加而增强; 水分过高则会阻塞土壤孔隙, 导致土壤呼吸强度减弱(周萍等, 2009).本研究期间0~10 cm土层深度土壤水分始终低于黄绵土田间持水量水平(焦菊英等, 2006), 未达到因土壤水分饱和而抑制土壤呼吸的状态(李洪建等, 2010).不同群落内, 土壤含水量变化过程存在差异:降雨补充的土壤水分在达乌里胡枝子群落中消减较快, 降雨后第5 d即回到对照水平, 而白羊草与铁杆蒿群落在降雨后7 d仍高于对照.这主要与群落盖度、光照及枯落物的持水能力有关.本试验中, 达乌里胡枝子群落盖度低于白羊草与铁杆蒿, 土壤水分易蒸发; 此外, 由于物种本身形态结构的差异, 白羊草和铁杆蒿枯落物的持水能力显著高于达乌里胡枝子(寇萌等, 2015).枯落物持水能力会影响枯落物分解, 持水能力较强的物种, 其分解过程相对较快, 表明土壤水分也是控制枯落物分解速率及其分解过程的重要因素, 这也将影响土壤呼吸的短期变化(Schnell et al., 1996).
 |
| 图 4 三种草地群落土壤呼吸速率和土壤含水量的关系 Fig. 4 Relationship between soil respiration and soil water content in three communities |
土壤呼吸对温度变化响应的敏感程度通常由Q10值表示.Q10值受土壤温度、土壤含水量、根生物量、枯落物等多种因素影响, 在不同群落和降雨量间均存在差异(Yuste et al., 2004).本试验结果表明, 降雨量对白羊草群落Q10值有显著影响, 且50 mm降雨处理显著高于15 mm降雨处理.50 mm降雨量下, 土壤温度能解释白羊草群落土壤呼吸59%~73%的变异, 表明在水分充足情况下, 温度是影响白羊草群落土壤呼吸的主导因子.达乌里胡枝子群落Q10值受月份影响较大, 7月土壤呼吸与土壤温度拟合的程度较6月显著, 可能与其地下生物量变化有关.研究表明, 7月达乌里胡枝子0~10 cm土层的地下生物量比6月有所下降(朱志诚等, 1994), 根呼吸的比重减少, 土壤微生物呼吸的组分增加, 因此对土壤温度的变化更为敏感.此外, 7月份群落内土壤含水量较高, 有利于提高微生物活性及其与呼吸底物的接触.研究认为, 土壤含水量的降低会导致Q10值降低(Qi et al., 2002), 本试验没有观测到土壤含水量与Q10值呈显著正相关的趋势, 但低土壤含水量下, 土壤呼吸与土壤温度的指数关系不显著, 仍可以反映Q10值对土壤水分存在一定的依赖性.3种草地群落土壤呼吸的Q10均值为1.56, 略低于黄土区其他草地生态系统的Q10值(1.74)(王爱国等, 2013).考虑到数据采自6月和7月气温较高的日间, 未包含低温月份的土壤呼吸速率, 可能会导致计算结果偏低, 因此, 有必要进行跨季节的长期监测, 以精确探明不同气象条件下土壤呼吸对温度的敏感性差异及其长期变化规律.
全球变暖背景下, 降雨格局改变将对干旱半干旱地区植被群落组成和结构产生重要影响(Báez et al., 2013).在退耕草地演替过程中, 年降水量的增加有利于以白羊草为代表的禾本科C4植物的生长与扩展(Liu et al., 2005b).据此我们推测, 随着降雨量的增加, 分布范围扩大的白羊草群落在短期降雨过程中将释放更多的CO2.因此, 适当考虑物种属性及其对降雨响应差异, 有助于准确估算降雨变化对陆地生态系统土壤呼吸和碳循环的效应.
5 结论(Conclusions)群落类型和降雨量均显著影响土壤含水量和土壤呼吸速率, 土壤温度在不同月份和降雨量下差异显著(p < 0.05).3种群落土壤呼吸对降雨的响应程度存在明显差异, 雨后土壤呼吸速率增量和平均土壤呼吸速率均以白羊草群落最高, 达乌里胡枝子群落最低.15 mm和50 mm降雨能够显著提高白羊草群落土壤呼吸速率, 铁杆蒿群落土壤呼吸速率仅在50 mm降雨后显著提升.土壤含水量和土壤呼吸速率呈显著对数函数关系(p < 0.05).3种群落Q10值为1.31~2.01, 其在不同月份和降雨量的差异表明, 物种属性和生长时期对Q10具有不同的影响方式.本试验重点关注6月和7月期间3种草地群落土壤呼吸随次降雨等级的动态变化, 有必要继续开展不同类型草地群落在不同生长阶段土壤呼吸对降雨的响应特征研究, 为评估降雨格局变化背景下黄土丘陵区退耕草地群落碳收支状况提供系统的基础数据.
Báez S, Collins S L, Pockman W T, et al. 2013. Effects of experimental rainfall manipulations on Chihuahuan Desert grassland and shrubland plant communities[J]. Oecologia, 172(4): 1117–1127.
DOI:10.1007/s00442-012-2552-0
|
Ball B C, Scott A, Parker J P. 1999. Field N2O, CO2 and CH4 fluxes in relation to tillage, compaction and soil quality in Scotland[J]. Soil & Tillage Research, 53(99): 29–39.
|
步秀芹, 徐学选, 康博文, 等. 2007. 黄土丘陵区铁杆蒿光合特性[J]. 林业科学, 2007, 43(9): 123–127.
|
Cable J M, Ogle K, Williams D G, et al. 2008. Soil texture drives responses of soil respiration to precipitation pulses in the Sonoran Desert:Implications for Climate Change[J]. Ecosystems, 11(6): 961–979.
DOI:10.1007/s10021-008-9172-x
|
Chen S, Lin G, Huang J, et al. 2008. Response of soil respiration to simulated precipitation pulses in semiarid steppe under different grazing regimes[J]. Journal of Plant Ecology, 1(4): 237–246.
DOI:10.1093/jpe/rtn020
|
Fu B, Chen L. 2000. Agricultural landscape spatial pattern analysis in the semi-arid hill area of the Loess Plateau, China[J]. Journal of Arid Environments, 44(3): 291–303.
DOI:10.1006/jare.1999.0600
|
Gestel M V, Merckx R, Vlassak K. 1993. Microbial biomass and activity in soils with fluctuating water contents[J]. Geoderma, 56(1/4): 617–626.
|
Gillon D, Houssard C, Joffre R. 1999. Using near-infrared reflectance spectroscopy to predict carbon, nitrogen and phosphorus content in heterogeneous plant material[J]. Oecologia, 118(2): 173–182.
DOI:10.1007/s004420050716
|
Grant R F, Rochette P. 1994. Soil Microbial respiration at different water potentials and temperatures:theory and mathematical modeling[J]. Soil Science Society of America Journal, 58(6): 1681–1690.
DOI:10.2136/sssaj1994.03615995005800060015x
|
黄懿梅, 安韶山, 刘连杰, 等. 2009. 黄土丘陵区土壤基础呼吸对草地植被恢复的响应及其影响因素[J]. 中国生态农业学报, 2009, 17(5): 862–869.
|
Huxman T E, Snyder K A, Tissue D, et al. 2004. Precipitation pulses and carbon fluxes in semiarid and arid ecosystems[J]. Oecologia, 141(2): 254–268.
DOI:10.1007/s00442-004-1682-4
|
焦菊英, 焦峰, 温仲明. 2006. 黄土丘陵沟壑区不同恢复方式下植物群落的土壤水分和养分特征[J]. 植物营养与肥料学报, 2006, 12(5): 667–674.
DOI:10.11674/zwyf.2006.0511 |
Jackson R B, Carpenter S R, Dahm C N, et al. 2001. Water in a changing world[J]. Ecological Applications, 11(4): 1027–1045.
DOI:10.1890/1051-0761(2001)011[1027:WIACW]2.0.CO;2
|
寇萌, 焦菊英, 尹秋龙, 等. 2015. 黄土丘陵沟壑区主要草种枯落物的持水能力与养分潜在归还能力[J]. 生态学报, 2015, 35(5): 1337–1349.
|
Knapp A K, Beier C, Briske D D, et al. 2008. Consequences of more extreme precipitation regimes for terrestrial ecosystems[J]. Bioscience, 58(9): 811–821.
DOI:10.1641/B580908
|
Knapp A K, Briggs J M, Koelliker J K. 2001. Frequency and extent of water limitation to primary production in a mesic temperate grassland[J]. Ecosystems, 4(1): 19–28.
DOI:10.1007/s100210000057
|
李国辉. 2010. 黄土丘陵区典型植物根际与非根际土壤性质差异的比较研究[D]. 杨凌: 西北农林科技大学
http://cdmd.cnki.com.cn/article/cdmd-10712-2010149356.htm |
李洪建, 严俊霞, 李君剑, 等. 2010. 黄土高原东部山区两种灌木群落的土壤碳通量研究[J]. 环境科学学报, 2010, 30(9): 1895–1904.
|
Liu B, Xu M, Henderson M, et al. 2005a. Observed trends of precipitation amount, frequency, and intensity in China, 1960–2000[J]. Journal of Geophysical Research Atmospheres, 110(D8): 211–211.
|
Liu W, Huang Y, An Z, et al. 2005b. Summer monsoon intensity controls C4/C3 plant abundance during the last 35 ka in the Chinese Loess Plateau:Carbon isotope evidence from bulk organic matter and individual leaf waxes[J]. Palaeogeography Palaeoclimatology Palaeoecology, 220(3/4): 243–254.
|
牛富荣, 徐炳成, 段东平, 等. 2011. 不同水肥条件下白羊草叶片叶绿素荧光特性研究[J]. 中国草地学报, 2011, 33(6): 75–81.
|
彭琴, 齐玉春, 董云社, 等. 2012. 干旱半干旱地区草地碳循环关键过程对降雨变化的响应[J]. 地理科学进展, 2012, 31(11): 1510–1518.
DOI:10.11820/dlkxjz.2012.11.012 |
Qi Y, Liu X, Dong Y, et al. 2014. Differential responses of short-term soil respiration dynamics to the experimental addition of nitrogen and water in the temperate semi-arid steppe of Inner Mongolia, China[J]. Journal of Environmental Sciences, 26(4): 834–845.
DOI:10.1016/S1001-0742(13)60509-4
|
Qi Y, Xu M, Wu J. 2002. Temperature sensitivity of soil respiration and its effects on ecosystem carbon budget:nonlinearity begets surprises[J]. Ecological Modelling, 153(1): 131–142.
|
Raich J W, Tufekciogul A. 2000. Vegetation and soil respiration:Correlations and controls[J]. Biogeochemistry, 48(1): 71–90.
DOI:10.1023/A:1006112000616
|
Rutter A J, Morton A J. 1977. A predictive model of rainfall interception in forests.Ⅲ.sensitivity of the model to stand parameters and meteorological variables[J]. Journal of Applied Ecology, 14(2): 567–588.
DOI:10.2307/2402568
|
Schlesinger W H, Andrews J A. 2000. Soil respiration and the global carbon cycle[J]. Biogeochemistry, 48(1): 7–20.
DOI:10.1023/A:1006247623877
|
Schnell S, King G M. 1996. Responses of methanotrophic activity in soils and cultures to water stress[J]. Applied & Environmental Microbiology, 62(9): 3203–3209.
|
孙智辉, 曹雪梅, 刘志超, 等. 2009. 陕西黄土高原地区降水日数和强度年代际变化[J]. 应用气象学报, 2009(4): 510–512.
|
唐龙, 梁宗锁, 杜峰, 等. 2006. 陕北黄土高原丘陵区撂荒演替及其过程中主要乡土牧草的确定与评价[J]. 生态学报, 2006, 26(4): 1165–1175.
|
Sponseller R. 2007. Precipitation pulses and soil CO2 flux in a Sonoran Desert ecosystem[J]. Global Change Biology, 13(2): 426–436.
DOI:10.1111/gcb.2007.13.issue-2
|
Tang J, Baldocchi D D. 2005. Spatial-temporal variation in soil respiration in an oak-grass savanna ecosystem in California and its partitioning into autotrophic and heterotrophic components[J]. Biogeochemistry, 73(1): 183–207.
DOI:10.1007/s10533-004-5889-6
|
Tian Y, Su D, Li F, et al. 2003. Effect of rainwater harvesting with ridge and furrow on yield of potato in semiarid areas[J]. Field Crops Research, 84(3): 385–391.
DOI:10.1016/S0378-4290(03)00118-7
|
Travis E H, Keirith A S, David T, et al. 2004. Precipitation pulses and carbon fluxes in semiarid and arid ecosystems[J]. Oecologia, 141(2): 254–268.
DOI:10.1007/s00442-004-1682-4
|
Xu M, Qi Y. 2001a. Spatial and seasonal variations of Q10 determined by soil respiration measurements at a Sierra Nevadan Forest[J]. Global Biogeochemical Cycles, 15(3): 687–696.
DOI:10.1029/2000GB001365
|
Xu M, Ye Q. 2001b. Soil-surface CO2 efflux and its spatial and temporal variations in a young ponderosa pine plantation in Northern California[J]. Global Change Biology, 7(6): 667–677.
DOI:10.1046/j.1354-1013.2001.00435.x
|
王爱国, 赵允格, 许明祥, 等. 2013. 黄土高原不同侵蚀区生物结皮土壤CO2通量日动态特征及其温度敏感性[J]. 环境科学学报, 2013, 33(9): 2605–2611.
|
王百群, 姜峻, 都全胜, 等. 2010. 黄土丘陵区人工草地牧草营养元素累积及土壤有机碳与养分特征[J]. 水土保持研究, 2010, 17(6): 127–132.
|
王会梅, 徐炳成, 李凤民, 等. 2005. 黄土丘陵区白羊草和柳枝稷适应性生长的比较[J]. 干旱地区农业研究, 2005, 23(5): 35–40.
|
温仲明, 焦峰, 刘宝元, 等. 2005. 黄土高原森林草原区退耕地植被自然恢复与土壤养分变化[J]. 应用生态学报, 2005, 16(11): 2025–2029.
DOI:10.3321/j.issn:1001-9332.2005.11.004 |
Yuste J C, Janssens I A, Carrara A R. 2004. Annual Q10 of soil respiration reflects plant phenological patterns as well as temperature sensitivity[J]. Global Change Biology, 10(2): 161–169.
DOI:10.1111/gcb.2004.10.issue-2
|
Zhang Y, Xu Y, Dong W, et al. 2006. A future climate scenario of regional changes in extreme climate events over China using the PRECIS climate model[J]. Geophysical Research Letters, 33(24): 194–199.
|
张海东, 汝海丽, 焦峰. 2015. 黄土丘陵区典型植被类型草本层生态学特性与环境解释[J]. 水土保持研究, 2015, 22(5): 100–105.
|
张娜, 梁一民. 2004. 黄土丘陵区天然草地地下/地上生物量的研究[J]. 草业学报, 2004, 11(2): 72–78.
|
周萍, 刘国彬, 薛萐. 2009. 草地生态系统土壤呼吸及其影响因素研究进展[J]. 草业学报, 2009, 18(2): 184–193.
DOI:10.11686/cyxb20090227 |
朱志诚, 贾东林. 1994. 达乌里胡枝子群落生物量初步研究[J]. 中国草地学报, 1994(3): 25–25.
|