 2017, Vol. 37
2017, Vol. 37



2. 岐阜大学 工学研究科, 日本岐阜市
2. Graduate School of Engineering, Gifu University, Gifu, Japan
蚯蚓堆肥作为一种简单易行安全可持续的处理技术而受到关注(Hartenstein et al., 1981; Castillo et al., 2013).蚯蚓堆肥稳定过程可以看作有机碳、氮向无机碳、氮的转化过程, 这个过程越彻底, 堆肥产物越稳定.前期实验发现, 蚯蚓堆肥稳定化过程大致可分为3个阶段, 即有机质降解阶段、氨化阶段和硝化阶段(陈学民等, 2016).蚯蚓堆肥过程中, 若氨化作用大于硝化作用, 易造成NH4+的积累(吴振斌等, 2004), 抑制脲酶活性(黄娟等, 2012), 影响尿素分解转化(关松荫编著, 1986).NH4+的积累影响基质pH稳定, 抑制硝化菌活性(贾程等, 2008; 孙洪伟等, 2015), 影响蚯蚓的生存环境, 增加了堆肥过程氮损失的趋势(贾程等, 2008; 单爱丽等, 2009).
硝化作用受很多因素的影响, 如pH、水分、氧气含量以及温度等(侯雪燕, 2014).将城镇污泥造粒后进行蚯蚓堆肥, 基质pH和水分相对稳定(Fu et al., 2016), 蚯蚓生物量、堆肥环境温度和基质好氧环境就成为影响堆肥过程中硝化作用的主要因素.研究蚯蚓生物量、堆肥环境温度和基质氧含量对硝化进程的影响, 有助于改进堆肥工艺条件, 消除基质产氨对蚯蚓的不利影响, 提高蚯蚓堆肥中氮素含量, 加速堆肥稳定化过程.
2 材料与方法(Materials and methods)污泥取自兰州市安宁区七里河污水处理厂脱水车间, 将取回的含水率80%左右的污泥制成5 mm和14.5 mm两种粒径作为实验用造粒污泥, 实验用反应器为ϕ36 cm×12 cm的不锈钢盆, 每个反应器中加4 kg实验用污泥, 供试蚯蚓为实验室喂养的赤子爱胜蚯蚓(Eisenia foetida), 设置生物量实验、温度实验和好氧条件实验.造粒污泥蚯蚓堆肥实验如图 1所示.
 |
| 图 1 蚯蚓堆肥实验 Fig. 1 Vermicomposting experiment |
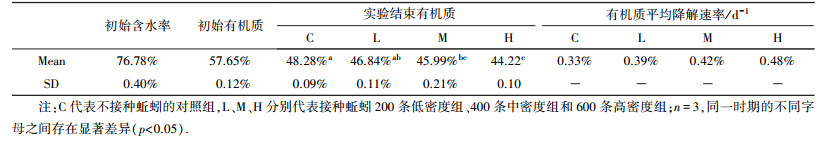
蚯蚓生物量实验设置无蚯蚓(对照组)、200条蚯蚓(低密度)、400条蚯蚓(中密度)、600条蚯蚓(高密度)4个密度组, 每个密度组设置3个重复, 共12个反应器.每个反应器中放5 mm粒径的实验用污泥4 kg.反应器覆盖塑料薄膜遮光保湿, 置于20 ℃恒温室, 每天人工翻动1次.前期每2 d取1次样, 16 d后每4 d取1次样, 实验进行28 d.
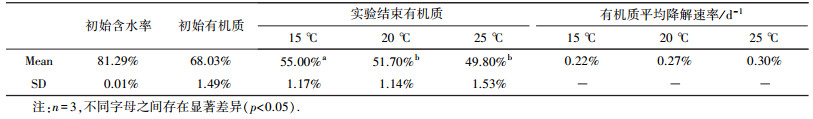
温度实验设置15、20、25 ℃ 3个温度, 每个温度设置3个重复, 共9个反应器.每个反应器中各加入5 mm粒径的实验用污泥4 kg和100条蚯蚓.反应器覆盖塑料薄膜遮光保湿, 堆肥环境温度分别为实验温度, 每天人工翻动1次.每隔10 d取1次样, 实验共进行60 d.
好氧条件实验分别取5 mm和14.5 mm两种粒径的污泥, 5 mm和14.5 mm粒径污泥各设3个重复, 每个反应器中加入实验用污泥4 kg, 接种200条蚯蚓.反应器覆盖塑料薄膜遮光保湿, 环境温度(20+0.1) ℃, 每天人工翻动1次.每隔10 d取1次样, 实验共进行60 d.
含水率由烘干重量法测定; 有机质采用灼烧法(550 ℃, 5 h)测定; NH4+采用氯化钾溶液提取分光光度法(HJ 634—2012);NO3-采用酚二磺酸比色法测定(鲍士旦主编, 2000).
分析平台为SPSS 19.0, 通过单因素方差分析(ANOVA)对同一指标不同处理组进行显著差异性分析(HSD), 处理组间差异显著性定义为p < 0.05.作图工具为Excel 2013.
3 结果与讨论(Results and discussion) 3.1 蚯蚓生物量的影响实验用污泥理化性质见表 1.
| 表 1 蚯蚓生物量实验污泥理化性质 Table 1 Physicochemical properties of the dewatered sludge with different earthworm biomasses |
由图 2a可以看出NH4+变化与蚯蚓生物量相关, 蚯蚓生物量越大NH4+含量越高, 在堆肥后期更加明显.图 2b中, 蚯蚓生物量对NO3-含量的影响前期无明显规律, 后期则与蚯蚓生物量显著正相关.
 |
| 图 2 不同蚯蚓生物量NH4+、NO3-随时间变化(同一时期的不同字母之间存在显著差异, p < 0.05) Fig. 2 Changes of NH4+ and NO3- during vermicomposting process with different earthworm biomasses |
蚯蚓的存在能够提高微生物活性(Hickman et al., 2008; Sen et al., 2009; Fu et al., 2016), 加速微生物对有机氮的利用, 在解聚和脱氨基作用下将有机氮转化成NH4+(赵彤等, 2014).另一方面, 蚯蚓代谢活动本身也会增加基质中NH4+含量(Aira et al., 2008; Gómez Brandón et al., 2010).堆肥前期污泥中微生物可利用的有机质丰富, 微生物转氨基作用产生的氨基态氮比蚯蚓代谢产NH4+占比高, 蚯蚓生物量对NH4+含量的影响并不明显.堆肥中期和后期, 基质中易利用营养物质逐渐减少, 微生物量也随之减少, 微生物产NH4+作用减弱, 基质中NH4+含量以蚯蚓自身代谢产NH4+为主, 这可能是堆肥中期和后期NH4+含量与蚯蚓生物量相关性更密切的原因.
NO3-与蚯蚓生物量相关性在堆肥后期趋于明显(图 2b), 这可能是因为堆肥前期易利用有机质含量高, 异养微生物新陈代谢旺盛, 堆肥总需氧量较高, 蚯蚓机械运动带入堆肥的氧量难以同时满足异养菌和硝化菌对氧的需求, 而异养菌获得氧的能力强于硝化菌(Laanbroek et al., 1994), 使硝化菌对氧的需求得不到满足, 从而活性受到了抑制.随着易利用有机质减少, 异养微生物需氧量也逐渐减少, 硝化菌获氧机会增加, 硝化作用也随之增强.同时堆肥过程中NH4+含量的升高, 增加了硝化底物浓度.而蚯蚓粪数量也会随堆肥进程而增加, 由此带来的硝化菌数量也不断增加(Parkin et al., 1999).这些因素的综合作用, 促进了后期污泥的硝化.
堆肥后期高密度组与中密度组的NH4+和NO3-含量之差高于其他密度组之间的差值, 说明NH4+和NO3-含量与蚯蚓生物量之间的相关性存在跃变, 即蚯蚓生物量达到某个密度之后, 其影响会更加显著.虽然NH4+的大量产生会对氨氧化菌和亚硝酸氧化菌活性产生抑制作用(贾程等, 2008; 孙洪伟等, 2015), 影响硝化速率, 但由图 2中NH4+和NO3-正相关可判断, 本实验中NH4+增加并未对硝化作用产生明显的抑制, 较高的NH4+反而提高了硝化反应底物浓度, 有利于硝化反应的进行, 这可能与蚯蚓粪中硝化菌的持续补充有关.
3.2 温度的影响温度是影响蚯蚓和微生物活性的重要因素之一, 也是堆肥工艺需要重点关注的.温度实验污泥理化性质见表 2.
| 表 2 温度实验污泥理化性质 Table 2 Physicochemical properties of the dewatered sludge with different temperatures |
温度实验蚯蚓堆肥过程NH4+变化见表 3.
| 表 3 温度实验蚯蚓堆肥过程NH4+变化 Table 3 Changes of NH4+ during vermicomposting process with different temperatures |

由表 3可知, 前30 d 15、20和25 ℃组之间NH4+含量并无显著差异, 第40 d 25 ℃组与15 ℃和20 ℃组NH4+含量开始出现显著差异性, 但15 ℃与20 ℃组之间NH4+含量并无显著差异.第50 d至实验结束3个温度组之间NH4+含量均出现显著差异.
通常, 氨化菌在35 ℃左右比较活跃(白帆等, 2011), 温度高的组氨化作用会更强烈, 但实验发现前30 d 3个温度组之间并未显现差异性.这可能是因为在城镇污泥蚯蚓堆肥过程中, 含氮有机物降解以及蚯蚓代谢产氨是NH4+的主要来源, 赤子爱胜蚓最适宜活动的温度范围为17~28℃(黄福珍著, 1982), 实验温度涵盖在其适宜活动的温度范围内, 因此可以推测温度对蚯蚓代谢活动影响不大.温度虽然会影响微生物代谢活性, 但由于堆肥前期微生物数量大活性相对较高, 可利用有机质含量高, 加之3个实验温度的温差仅有10 ℃, 温度的影响极为有限.虽然3个温度组产NH4+能力25 ℃>20 ℃>15 ℃, 但远没有达到显著性水平.随着堆肥进程的进行, 由温差引起的NH4+增量累积使25 ℃组与15 ℃和20 ℃组之间在40 d首先达到了显著性水平.50 d时, 3个温度组NH4+含量差异均达到显著性水平.可见温度对微生物和蚯蚓产NH4+的影响是一个渐变累积过程, 温度高更有利于这一累积过程产生量的跃变.
从图 3所示NO3-变化可知, 前40 d NO3-并未因温度的不同而出现显著差异, 40 d后不同温度NO3-含量出现显著差异.25 ℃组NO3-增量远大于20 ℃组与15 ℃组, 表明温度对堆肥后期硝化作用影响显著, 并且这种影响在不同的温度范围内显著不同.这可能是因为40 d后堆肥积累了足够量的硝化底物NH4+, 同时硝化菌获氧环境向着更有利于硝化菌活性的方向发展, 使硝化进程得以启动.但25 ℃处于硝化菌的适中温度(李明峰等, 2014), 酶活性相对较高, 加之25 ℃组积累的底物NH4+浓度高于20 ℃组与15 ℃组, 因此25 ℃组NO3-含量远高于20 ℃组与15 ℃组.图 3所示NO3-显著变化均发生在第40 d后, 表明在15~25℃范围内提高温度对蚯蚓堆肥硝化进程的影响较小, 但对硝化强度影响较大.
 |
| 图 3 不同温度下NO3-随时间变化(同一时期的不同字母之间存在显著差异, p < 0.05) Fig. 3 Changes of NO3- during vermicomposting process with different temperatures |
氧是好氧微生物功能能否得以充分发挥的重要因素, 城镇污泥有机质含量丰富, 微生物活性高, 充足的氧量是保障有机质快速降解的必要条件.脱水后污泥流动性差, 氧向污泥内部扩散阻力大, 通常仅表面较薄的污泥层处于好氧状态, 改变脱水污泥块的大小可以调整污泥好氧区和非好氧区体积比值.无论污泥粒径大小如何, 好氧层厚度基本相同, 因此污泥粒径越小, 污泥好氧体积占比就越大.若以M1、M2分别表示小粒径污泥、大粒径污泥好氧区与非好氧区体积比值, 则一定有M1 > M2.不同M值实验污泥理化性质见表 4.
| 表 4 不同M值实验污泥理化性质 Table 4 Physicochemical properties of the dewatered sludge with different M values |
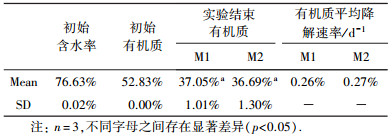
图 4a中M1, M2的NH4+的变化趋势基本一致, 表明氨化作用与M值大小关系并不十分密切, 这可能是因为脱氨基作用在好氧和厌氧条件下均能进行的缘故(顾夏声等, 2011).
 |
| 图 4 不同M值NH4+、NO3-随时间变化(*表示不同处理之间存在显著差异, p<0.05) Fig. 4 Changes of NH4+ and NO3- during vermicomposting process with different M values |
图 4b可以发现, M1的硝化始于第20 d, 而M2的硝化则始于第40 d, 这表明较高的M值有助于污泥硝化进程的提前, 这可能与充足的氧量更有利于自养硝化菌的硝化作用有关(Laanbroek et al., 1994).由于供氧充分条件下的氨氧化菌种类和数量明显多于供氧不充分条件下的(孙志远, 2012), 因此颗粒污泥中硝化细菌仅存在于污泥外层300 μm内的含氧区(Jang et al., 2003).可见, 提高M值能够改善污泥与氧接触的面积, 增加硝化菌的获氧机会, 使硝化更易进行.
4 结论(Conclusions)蚯蚓生物量对氨化和硝化影响随堆肥时间逐渐增强.氨化与硝化强度与蚯蚓生物量之间的相关性存在跃变, 当蚯蚓生物量大于一定值后蚯蚓的影响会更加显著.
温度对微生物和蚯蚓产NH4+的影响是一个渐变累积过程, 温度高更有利于这一累积过程产生量的跃变.15~25℃范围内提高温度对蚯蚓堆肥硝化进程的影响较小, 对硝化强度影响较大.越接近蚯蚓适宜生长的环境温度, 温度对氨化和硝化作用的影响就越大.
蚯蚓堆肥过程氧含量对氨化过程影响极小, 但对硝化过程影响显著.提高堆肥污泥与氧的接触面积, 有利于提前污泥硝化进程, 减少NH4+的积累量.
Aira M, Sampedro L, Monroy F, et al. 2008. Detritivorous earthworms directly modify the structure, thus altering the functioning of a microdecomposer food web[J]. Soil Biology and Biochemistry, 40(10): 2511–2516.
DOI:10.1016/j.soilbio.2008.06.010
|
白帆, 王晓昌. 2011. 人粪便好氧堆肥温度对氮迁移转化的影响[J]. 环境化学, 2011, 30(7): 1266–1270.
|
鲍士旦. 2000. 土壤农化分析[M]. 北京: 中国农业出版社: 50–53.
|
Castillo J M, Romero E, Nogales R. 2013. Dynamics of microbial communities related to biochemical parameters during vermicomposting and maturation of agroindustrial lignocellulose wastes[J]. Bioresource Technology, 146: 345–354.
DOI:10.1016/j.biortech.2013.07.093
|
陈学民, 雷旭阳, 伏小勇, 等. 2016. 温度对蚯蚓处理城镇污泥稳定化过程的影响[J]. 环境科学学报, 2016, 36(6): 2079–2084.
|
Fu X, Cui G, Huang K, et al. 2016. Earthworms facilitate the stabilization of pelletized dewatered sludge through shaping microbial biomass and activity and community[J]. Environmental Science and Pollution Research, 23(5): 4522–4530.
DOI:10.1007/s11356-015-5659-0
|
Gómez-Brandón M, Lazcano C, Lores M, et al. 2010. Detritivorous earthworms modify microbial community structure and accelerate plant residue decomposition[J]. Applied Soil Ecology, 44(3): 237–244.
DOI:10.1016/j.apsoil.2009.12.010
|
关松荫编著. 1986. 土壤酶及其研究法[M]. 北京: 农业出版社. 294
|
顾夏声, 胡洪营, 文湘华, 等. 2011. 水处理生物学(第5版)[M]. 北京: 中国建筑工业出版社: 218.
|
Hartenstein R, Hartenstein F. 1981. Physicochemical changes effected in activated sludge by the earthworm eisenia foetida1[J]. Journal of Environmental Quality, 10(3): 377–381.
|
Hickman Z A, Reid B J. 2008. Earthworm assisted bioremediation of organic contaminants[J]. Environment International, 34(7): 1072–1081.
DOI:10.1016/j.envint.2008.02.013
|
侯雪燕. 2014. 土壤pH对硝化作用和氨氧化微生物群落结构的影响[D]. 重庆: 西南大学. 3
http://cdmd.cnki.com.cn/article/cdmd-10635-1014261588.htm |
黄福珍. 1982. 蚯蚓[M]. 北京: 农业出版社: 44.
|
黄娟, 李稹, 张健. 2012. 改良靛酚蓝比色法测土壤脲酶活性[J]. 土木建筑与环境工程, 2012, 34(1): 102–107.
|
Jang A, Yoon Y H, Kim I S, et al. 2003. Characterization and evaluation of aerobic granules in sequencing batch reactor[J]. Journal of Biotechnology, 105(1): 71–82.
|
贾程, 张增强, 张永涛. 2008. 污泥堆肥过程中氮素形态的变化[J]. 环境科学学报, 2008, 28(11): 2269–2276.
DOI:10.3321/j.issn:0253-2468.2008.11.016 |
Laanbroek H J, Bodelier P L E, Gerards S. 1994. Oxygen consumption kinetics of Nitrosomonas europaea and Nitrobacter hamburgensis grown in mixed continuous cultures at different oxygen concentrations[J]. Archives of Microbiology, 161(2): 156–162.
DOI:10.1007/BF00276477
|
李明峰, 马闯, 赵继红, 等. 2014. 污泥堆肥臭气的产生特征及防控措施[J]. 环境工程, 2014(1): 92–96.
|
Parkin T B, Berry E C. 1999. Microbial nitrogen transformations in earthworm burrows[J]. Soil Biology and Biochemistry, 31(13): 1765–1771.
DOI:10.1016/S0038-0717(99)00085-1
|
Sen B, Chandra T S. 2009. Do earthworms affect dynamics of functional response and genetic structure of microbial community in a lab-scale composting system?[J]. Bioresource Technology, 100(2): 804–811.
DOI:10.1016/j.biortech.2008.07.047
|
单爱丽, 王帆, 严红. 2009. 城市生活垃圾好氧堆肥性质变化的研究[J]. 大连大学学报, 2009(3): 57–59.
|
孙洪伟, 尤永军, 白宝霞, 等. 2015. 游离氨对硝化菌活性的短期抑制影响试验研究[J]. 中国给水排水, 2015, 31(3): 19–23.
|
孙志远. 2012. 堆肥化过程中氨氧化菌群多样性分析[D]. 大庆: 黑龙江八一农垦大学. 41
http://cdmd.cnki.com.cn/article/cdmd-10223-1012389562.htm |
吴振斌, 徐光来, 周培疆, 等. 2004. 复合垂直流人工湿地对不同氮污水的净化[J]. 环境科学与技术, 2004, 27(S1): 30–32.
|
赵彤, 蒋跃利, 闫浩, 等. 2014. 土壤氨化过程中微生物作用研究进展[J]. 应用与环境生物学报, 2014(2): 315–321.
|