 2018, Vol. 38
2018, Vol. 38



2. 南开大学, 天津市生物质固废资源化技术工程中心, 天津 300071
2. Tianjin Engineering Research Center of Biomass Solid Waste Resources Technology, Nankai University, Jinnan District, Tianjin 300071
随着人口增多、工业化进程深入,我国土壤污染问题日益严重,耕地土壤环境质量尤为堪忧,主要表现为氮素淋失、持久性有机污染物和重金属含量超标(环境保护部等, 2014).一方面,植物的根部吸收土壤中的持久性有机污染物和重金属,对农作物生长及产量造成不良影响(程新伟, 2011),例如,我国因土壤重金属污染造成的粮食减产达1.0×105 t,造成经济损失2.0×1010元(罗小玲等, 2014);另一方面,污染物可以通过地表径流、淋溶、挥发等途径(葛鑫等, 2003)进入水体和大气,不仅造成水质恶化和大气污染,还可进一步通过食物链进入人体,危害身体健康(赵玲等, 2017).因此,亟需寻找高效的手段治理土壤污染.
针对土壤污染问题,主要的土壤修复技术有热处理、固定-稳定化、淋洗、氧化-还原、光催化降解、植物修复和微生物修复等.这些修复技术中,一部分操作复杂、能耗大、处理费用高,另一部分易产生二次污染(王连超, 2017).而吸附法因效果显著、成本低廉、易于操作等特点,在治理重金属、有机物污染等方面存在优势(朱银涛等, 2018).自Lehmann(2007)提出亚马逊黑土的功效,学者们发现,利用农林废弃物缺氧热解生成的生物炭是一种孔隙结构发达、比表面积大、含氧官能团丰富、具备优良吸附性能的材料(Ahmad et al., 2014).因此,为获得吸附性能优越的生物炭,学者们致力于探究生物质原料、热解温度、改性条件对生物炭理化性质的影响.金属离子或金属氧化物的改性,可以使金属离子与污染物发生离子交换,从而增强生物炭对污染物的修复效果.有学者(孟繁健等, 2017)通过对酸改性生物炭负载零价铁来研究其对六价铬Cr(Ⅵ)的修复,发现生物炭促进Cr(Ⅵ)还原及总Cr向残渣态转化;也有经氢氧化钠、氯化钙、高锰酸钾、三氯化铁、二氧化锰改性,探究生物炭对镉、砷、多环芳烃的修复效果(赵明静等, 2016; 董双快等, 2016),但这类研究仍不够系统.
我国小麦、玉米秸秆产量巨大,占秸秆总量的50%以上,是制备生物炭的常见原料(曹志宏等, 2018).但不同秸秆种类和热解温度均会造成热解过程的差异,导致生物炭理化性质的改变,从而对生物炭的吸附效果产生影响(薛俊杰, 2016).因此,本文以小麦秸秆和玉米秸秆为生物质原料,探讨热解温度和改性对生物炭理化性质的影响;测定生物炭对氨氮、铅离子(Pb(Ⅱ))和乙草胺的饱和吸附量,选择吸附效果最佳的生物炭,研究其对氨氮、Pb(Ⅱ)和乙草胺的吸附动力学、热力学和吸附机理,以期对修复土壤污染提供理论依据.
2 材料与方法(Materials and methods) 2.1 生物炭的制备及表征将小麦和玉米秸秆(取自天津郊区农田)用超纯水(18.25 MΩ·cm)洗涤数次,于105 ℃下烘干至恒重,粉碎后(40目)称取20 g于氮气气氛下进行裂解(升温速率5 ℃· min-1,恒温2 h),热解温度分别为300、400、500、600、700 ℃,冷却至室温后,粉碎至80目,小麦/玉米秸秆生物炭编号为XB/YB-温度.
将生物炭与无CO2的超纯水/氯化钙(CaCl2,本实验所用试剂均为分析纯,购自科密欧化学试剂有限公司)溶液混合后用pH计(pH400,alalis)测定;采用元素分析仪(EURO EA3000,利曼)分析样品的C、H和N含量;用高温灰化法(800 ℃,恒温3 h)测定灰分含量,并通过质量平衡法计算O含量;生物炭的比表面积采用比表面积快速分析仪(ASAP 2460,micromeiritics)测定;采用氯化钡-硫酸强迫交换法测定阳离子交换量(CEC);利用傅立叶变换红外光谱仪(IS50,Thermofisher)按KBr压片法测定4000~400 cm-1的红外光谱;利用X射线衍射仪(Ulitma Ⅳ,日本理学)表征材料的晶相结构.
2.2 生物炭的改性将0.01 mol· L-1的氯化镁(MgCl2·6H2O)溶液以1:50(m:V)的比例加入分别含有小麦秸秆、玉米秸秆及其生物炭样品的100 mL三角瓶,强力搅拌3 h后过滤、洗涤、烘干,即制得MgCl2改性生物炭(秸秆改性样品需绝氧热裂解);分别配制相同浓度的乙酸镁(Mg(CH3COO)2·4H2O)、硝酸镁(Mg(NO3)2)溶液,重复上述操作.样品编号(以Mg(NO3)2溶液为例):小麦/玉米秸秆生物炭改性样品为XB/YB-Mg(NO3)2,小麦/玉米秸秆改性生物炭样品为XM/YM-Mg(NO3)2.
2.3 吸附实验饱和吸附量试验:将准确称取的0.0600 g吸附性能优越的改性生物炭样品与30.00 mL氯化铵(NH4Cl)溶液(25.00 mg· L-1)置于三角瓶中,放置空气摇床内恒温(25 ℃)振荡1440 min,然后用0.45 μm滤膜过滤后采用纳氏试剂分光光度法测定滤液氨氮浓度.分别加入同体积硝酸铅(Pb(NO3)2)溶液(25.00 mg· L-1)、乙草胺溶液(10 mg· L-1)重复上述操作.Pb(Ⅱ)和乙草胺浓度分别采用原子吸收光谱仪(contrAA 700,耶拿)和高效液相色谱仪(Thermo,UltiMate 3000)进行测定,3种吸附体系中Mg(Ⅱ)浓度利用原子吸收光谱仪测定.每组试验设置3组平行试验.
吸附动力学试验:将准确称取的0.0600 g吸附性能优越的改性生物炭样品与30.00 mL NH4Cl溶液(25.00 mg· L-1)置于三角瓶中,放置空气摇床内恒温(25 ℃)振荡一定时间(5、10、30、45、60、120、240、480、960、1440 min),然后用0.45 μm滤膜过滤后测定氨氮浓度;分别加入同体积Pb(NO3)2溶液(25.00 mg· L-1)、乙草胺溶液(10 mg· L-1)重复上述操作并测定Pb(Ⅱ)和乙草胺浓度.每组试验设置3组平行试验.
吸附热力学试验:将准确称取的0.0600 g吸附性能优越的改性生物炭样品与30.00 mL不同浓度的NH4Cl溶液(5.00、10.00、25.00、40.00、50.00 mg· L-1)置于三角瓶中,放置恒温空气摇床内于不同温度下(15、25、35 ℃)振荡1440 min,然后用0.45 μm滤膜过滤后测定氨氮浓度;分别加入同体积不同浓度Pb(NO3)2溶液(5.00、25.00、50.00、100.00 mg· L-1)、乙草胺溶液(5.00、10.00、50.00 mg· L-1)重复上述操作并测定Pb(Ⅱ)和乙草胺浓度.每组试验设置3组平行试验.
2.4 数据分析吸附量计算:
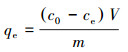
|
(1) |
式中,qe为吸附结束时单位质量吸附剂的吸附量(mg·g-1);c0和ce分别为溶液中吸附质的初始浓度和吸附平衡时的浓度(mg· L-1);V为溶液体积(mL);m为生物炭的投加量(g).
吸附动力学:准一级吸附动力学方程假定吸附速率受扩散步骤控制,描述的是物理吸附过程,其表达式为(Lagergren, 1898):

|
(2) |
进行积分后可得到公式:
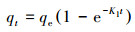
|
(3) |
式中,qt和qe分别为吸附时间t、吸附平衡时,吸附剂对吸附质的吸附量(mg·g-1);K1为吸附质在吸附剂表面上吸附的准一级速率常数h-1(或min-1).
准二级吸附动力学方程认为吸附的本质是化学吸附,吸附的实质在于体系内各物质间的电子转移或共用.其表达式(Zhang et al., 2018)为:

|
(4) |
吸附等温线:Langmuir型吸附等温式视吸附剂表面为均一化表面,吸附行为为单分子层吸附,吸附平衡时的吸附量为饱和吸附量.其中平衡吸附量qe与液相平衡浓度ce的关系表达式为:
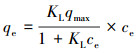
|
(5) |
式中,qmax为特定吸附条件下吸附剂的最大吸附量(mg·g-1);KL为Langmuir吸附平衡常数(L·mg-1).
其线性表达式为:

|
(6) |
Freundlich型吸附等温式为经验性公式,但对吸附剂表面不均一、吸附为复杂的等温吸附可进行很好的拟合.
平衡吸附量qe与液相平衡浓度ce的关系表达式为:
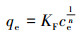
|
(7) |
式中,qe为吸附平衡时的吸附量(mg·g-1);KF为Freundlich吸附平衡常数;n为常数,与吸附剂对吸附质的亲和力有关.
对该式求对数后,其线性表达式为:
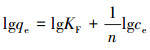
|
(8) |
吸附热力学:吸附热力学焓变可以更直观地反映吸附剂与吸附质分子之间以及吸附剂与溶剂之间的作用,能确定吸附是吸热还是放热、是否是自发的过程.表达式为:
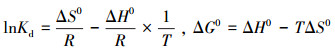
|
(9) |
式中,Kd为分配系数(mL·g-1);T为绝对温度(K);R为理想气体常数,8.3145 J·moL-1·K-1;ΔG0为吉布斯自由能变(kJ·moL-1);ΔH0为焓变(kJ·moL-1);ΔS0为熵变(J·moL-1·K-1).
3 结果与讨论(Results and discussion) 3.1 生物炭的基本特征及理化性质 3.1.1 生物炭的基本性质由表 1可知,总体而言,小麦秸秆生物炭的灰分、pH(CaCl2/H2O)、比表面积、总孔容及CEC含量均高于玉米秸秆生物炭.当裂解温度小于700 ℃时,随着温度升高,小麦和玉米秸秆生物炭的灰分含量、CEC、pH(CaCl2/H2O)、比表面积及总孔容升高,产率降低;裂解温度为700 ℃时,pH(CaCl2/H2O)、比表面积及总孔容降低.由此推断,由于裂解温度高,孔隙结构逐渐发育,微孔增多,造成比表面积增大.但过高的温度使得微孔结构坍塌,因此,700 ℃样品比表面积减小.同时,随着裂解温度升高,碳化程度增大,导致灰分含量高、生物炭残留量低,最终造成产率下降.灰分中的矿质元素如Na、K、Mg、Ca等使得生物炭CEC含量升高,且矿质元素以氧化物或碳酸盐形式存在,溶于水后呈碱性(Yuan et al., 2011; 马锋锋等, 2017a),则灰分越高,CEC越高、pH越高.根据元素组成可知,H/C、O/C、(O+N)/C大致随温度升高而降低.其中,H/C可表征生物炭的芳香性,H/C值越小表示芳香性越强,生物炭结构越稳定.O/C和(O+N)/C可代表生物炭的亲水性和极性,O/C和(O+N)/C值越高说明亲水性和极性越强(Chen et al., 2010).由此推断,在600 ℃以内,随着温度升高,生物炭样品的芳香性更强、结构更加稳定、亲水性和极性逐渐减弱,且小麦秸秆生物炭芳香性强于玉米秸秆生物炭.
| 表 1 生物炭的基本理化性质 Table 1 The basic physicochemical properties of biochars |
将改性生物炭与原始生物炭(XB/YB-500)的理化性质进行对比,得出结论:改性生物炭样品的O/C和(O+N)/C值增大,即改性生物炭的极性和亲水性增强.灰分含量、pH均增大,且CEC是原始生物炭的10倍左右.有文献(Lee et al., 2010; 杨惟薇, 2014)指出生物炭的O/C与CEC呈正相关,这与改性生物炭的CEC远高于原始样品的结果一致.考虑是由于Mg(Ⅱ)负载在生物炭表面,导致CEC含量升高.改性生物炭样品的比表面积、总孔容减小,可能是Mg(Ⅱ)的负载占据生物炭的孔隙,同时降低造孔能力造成的(Fang et al., 2014).
3.1.2 XRD分析由图 1可知,不同热解温度下制备的生物炭样品的XRD图谱存在一定的差异.一般认为,在2θ=15°~22°出现的宽缓弥散峰是由于脂肪链引起的(闵凡飞等, 2006).样品XB-300在2θ=26.5°、29.3°处为石英衍射峰,衍射峰强度均随热解温度升高而增大,由无定型变为晶型SiO2(Zhang et al., 2018; Cao et al., 2010);2θ=31.5°处为碳酸钙(006)晶面的衍射峰,衍射峰强度随热解温度升高而减小,考虑是温度升高使得碳酸盐分解造成的.
 |
| 图 1 不同热解温度下制备生物炭的X射线衍射光谱图 Fig. 1 X-ray diffraction spectra of biochar prepared under different pyrolysis temperatures |
图 1a, 1 b在2θ约为28.5°、40.6°、50.5°时均有明显的钾盐衍射峰,该衍射峰随着温度升高强度增大(何莉莉, 2013),且玉米秸秆生物炭钾盐峰强度更大.样品YB-700在此处的衍射峰强度最大,考虑是因为温度升高,其他物质逐步分解造成的.
利用XRD对3种经镁改性的生物炭样品与未改性样品进行比较,其结果如图 1c, 1d所示.镁改性生物炭均在2θ=42.7°有明显衍射峰,该峰应为MgO的特征峰,证明将镁离子已负载在生物炭上.
3.1.3 傅里叶变换红外光谱图由图 2a可知,热解温度不同,生物炭表面官能团的种类和数量不尽相同.样品XB-300在3100~3600 cm-1处峰型较宽,之后随着温度升高,小麦秸秆生物炭于3422 cm-1处出峰,该峰为羟基(—OH)的伸缩振动(李洋等, 2015),说明高温使小麦秸秆的—OH增多.XB-300和XB-400在2929 cm-1处的振动峰为脂肪性—CH2—不对称伸缩振;1701 cm-1处为酯类(C══O)的特征吸收峰,随着制备温度升高,这两种峰逐渐消失(马锋锋等, 2017b).1609 cm-1处的峰为芳环C══C的伸缩振动,随着裂解温度升高,该峰出现红移(Chun et al., 2004).1432 cm-1处的峰为羧基(—COOH)和醛基(—CHO)(Chun et al., 2004).1105 cm-1处的峰为纤维素或半纤维素的C—O—C振动(Keiluweit et al., 2010).876、796、619 cm-1处的峰为C—H面外弯曲振动,且随温度升高,振动峰增强(Chun et al., 2004).
 |
| 图 2 不同热解温度下制备生物炭的红外光谱图 Fig. 2 Infrared spectra of biochar prepared under different pyrolysis temperatures |
由图 2b可知,玉米秸秆生物炭在3100~3600 cm-1处峰型较宽,为—OH的伸缩振动,且随制备温度升高,该振动峰逐渐减弱,这是由于结合水的脱离导致氢键结合的—OH逐渐断裂(马锋锋等, 2017c).2927 cm-1处的吸收峰为脂肪性—CH2—不对称伸缩振动(Yuan et al., 2011; Wang et al., 2008),随温度升高,振动峰减弱,说明高温使玉米秸秆的脂肪性烷基链数量减少.1702 cm-1处是—COOH中C══O的伸缩振动峰;1570~1610 cm-1处的吸收峰为芳环C══O、C══C的伸缩振动,温度升高后该峰逐渐消失,表明玉米秸秆生物炭中纤维素含量逐渐降低(马锋锋等, 2017a).1442 cm-1处的吸收峰为C══O键伸缩振动(Chen et al., 2010; Wu et al., 2012),1259 cm-1处的吸收峰为酚羟基的伸缩振动吸收峰(马锋锋等, 2017c).1068 cm-1处为C—O的伸缩振动峰,649~878 cm-1为C—H的伸缩振动.由此可见,两种秸秆的生物炭样品所含官能团种类基本一致,但小麦秸秆生物炭的特征峰强度更大,即官能团数量多于玉米秸秆.
由图 2c, 2d可知,改性前后生物炭表面官能团种类基本无变化,但部分吸收峰强度增大.小麦秸秆改性样品在1701 cm-1处的C══O吸收峰增强;玉米秸秆改性样品的—COOH、C══O、C══C、C—O、—OH特征峰强度均有增大.Mg(CH3COO)2、MgCl2、Mg(NO3)2改性生物炭的含氧官能团数量增大,则极性增强,这与改性生物炭O/C值增大的结论一致.
3.2 吸附实验结果 3.2.1 饱和吸附量结果原始玉米和小麦秸秆生物炭对氨氮的吸附容量分别为1.63和1.54 mg·g-1,由图 3a可知,改性样品XM-Mg(CH3COO)2对氨氮的吸附容量较大,为6.57 mg·g-1,是原始小麦生物炭样品吸附量的4.27倍.改性生物炭样品XM/YM-Mg*系列对氨氮的吸附效果略优于XB/YB-Mg*系列,说明秸秆经镁溶液改性后制备的生物炭样品的改性效果最好.由图 3b可明显看出,改性样品XM-Mg(CH3COO)2对Pb(Ⅱ)的吸附容量最大,为16.81 mg·g-1,是原始生物炭样品吸附量的8.24倍,改性效果显著.图 3c中改性样品XM-Mg(CH3COO)2对乙草胺的吸附容量最大,为9.79 mg·g-1,是原始生物炭样品吸附量的4.78倍;因此选择XM-Mg(CH3COO)2样品进行后续吸附实验.
 |
| 图 3 生物炭样品对3种吸附质的吸附效果 Fig. 3 Adsorption of three pollutants by biochar samples |
由图 4a可知,样品XM-Mg(CH3COO)2对氨氮、Pb(Ⅱ)和乙草胺的吸附在初始阶段迅速上升,对氨氮和乙草胺的吸附在5 min内迅速升高,之后吸附量持续增大,但逐步趋于平缓;Pb(Ⅱ)的吸附平衡时间较长,240 min内吸附量显著升高,之后趋于平缓.因此,根据吸附质种类,将吸附时间控制在较短范围内,可提高吸附效率、降低能耗.对吸附过程进行准一级和准二级吸附动力学方程拟合,由图 4b和表 2可知,该样品对3种吸附质的吸附均符合准二级吸附动力学,相关系数R2均在0.999以上.实验测得的最大吸附量与准二级动力学计算的平衡吸附量基本一致,由此判断吸附过程以化学吸附为主.
 |
| 图 4 生物炭对3种吸附质的吸附动力学拟合曲线 Fig. 4 Adsorption kinetics of three pollutants on biochar |
| 表 2 生物炭对3种吸附质的吸附动力学参数 Table 2 Kinetics parameters of three pollutants by biochar |
图 5a、5b、5c分别为不同温度下,样品XM-Mg(CH3COO)2对3种吸附质的Langmuir模型的等温拟合曲线,图 5d、5e、5f为Freundlich模型的等温拟合曲线,拟合参数如表 3所示.根据R2推断,生物炭样品对氨氮的吸附过程符合Langmuir模型.该样品对Pb(Ⅱ)的吸附量随温度升高而升高,说明较高温度下Pb(Ⅱ)的吸附效率高,且该吸附过程符合Freundlich模型,认为以多层吸附为主.该样品对乙草胺的吸附量随温度升高而升高,该吸附过程对两种模型都符合,但Langmuir模型的相关系数略高,认为以单分子层均一吸附为主.
 |
| 图 5 生物炭对3种吸附质的吸附等温拟合曲线 Fig. 5 Adsorption isotherms of three pollutants on biochar |
| 表 3 生物炭对/3种吸附质的吸附等温方程拟合参数 Table 3 Isotherm parameters of three pollutants sorption by biochar |
图 6a、6b、6c分别为生物炭对氨氮、Pb(Ⅱ)、乙草胺的吸附热力学拟合曲线.由表 4可知,生物炭对氨氮的吸附过程中,ΔH0、ΔG0均为负值,说明该吸附过程是放热自发的.ΔS0随着氨氮初始浓度的增大,由负到正,说明吸附由有序变得无序.
 |
| 图 6 生物炭对3种吸附质的吸附热力学拟合曲线 Fig. 6 Adsorption thermodynamics of three pollutants on biochar |
| 表 4 生物炭对3种吸附质的热力学参数 Table 4 Thermodynamic parameters of three pollutants sorption by biochar |
生物炭对Pb(Ⅱ)的吸附过程中,ΔH0均为正值,且随温度升高吸附量增大,说明该吸附过程是吸热的;ΔS0随着初始浓度的增加呈现出先升后降的趋势且均为正值,说明无序度先增后降;在Pb(Ⅱ)初始浓度较低时,ΔG0为负值,说明吸附可自发进行;随着初始浓度增大,ΔG0增大为正值,但随着温度的升高,在308.15 K下ΔG0降低为负值,可自发进行.因此推断,在较高温度和适宜Pb(Ⅱ)浓度的条件下对Pb(Ⅱ)的吸附效果更好.
生物炭对乙草胺的吸附过程中,ΔH0、ΔS0均为正值,说明该吸附过程是无序吸热的;当乙草胺初始浓度为5、10 mg· L-1时,ΔG0 < 0,说明可以自发进行,但当浓度增大至50 mg· L-1时,ΔG0为正值,且随着温度的升高,ΔG0逐渐降低.
3.3 吸附机理一般而言,吸附剂的比表面积越大,越有利于对吸附质的表面吸附作用.但改性生物炭的比表面积降低并未造成吸附效果的降低,说明表面吸附并不是主要的吸附机制.小麦秸秆改性样品的官能团数量小于玉米秸秆改性样品,但吸附效果优于玉米秸秆改性样品,说明官能团络合亦不是主要的吸附机制.
分别测定不同时间段氨氮、Pb(Ⅱ)和乙草胺吸附体系中Mg(Ⅱ)的浓度,并计算其离子交换百分比,公式如下(以Pb(Ⅱ)吸附体系为例):
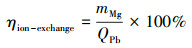
|
(10) |
式中,mMg表示生物炭释放Mg(Ⅱ)的量,QPb表示生物炭对Pb(Ⅱ)的吸附量.
由图 7可知,氨氮吸附体系中的Mg(Ⅱ)浓度高于空白对照组,且其整个吸附体系内的平均离子交换百分比为42.98%.因此认为改性生物炭对氨氮的吸附以离子交换为主,这与文献(郁强强, 2015)结论一致.
 |
| 图 7 各吸附体系中Mg(Ⅱ)浓度的变化趋势 Fig. 7 The variation trend of Mg(Ⅱ) concentration in each adsorption system |
生物炭表面官能团对氨氮的固定虽不是主要的吸附机制,但也有一定的吸附效果(丁绍兰等, 2010; 谢淘, 2015).因此,生物炭对氨氮的吸附机理如下所示:
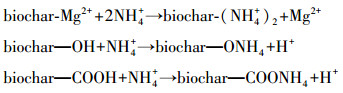
|
(11) |
随着吸附时间的增长,Pb(Ⅱ)吸附体系中的Mg(Ⅱ)浓度逐渐升高.与对照组相比,Pb(Ⅱ)的存在对Mg(Ⅱ)浓度影响显著.整个吸附体系内的平均离子交换百分比为44.79%,因此, 改性生物炭对Pb(Ⅱ)的作用机理与氨氮吸附体系类似,将Pb(Ⅱ)吸附在生物炭表面,而负载在生物炭上的Mg(Ⅱ)交换到土壤中.该改性方式不仅可吸附土壤中的重金属离子,还将营养元素释放到土壤中,提升土壤肥力.
生物炭对有机物的吸附主要以表面吸附、分配作用、π-π电子供体受体、表面含氧官能团、静电作用等机制为主.因此,乙草胺吸附体系中Mg(Ⅱ)浓度与空白对照组差异不大.改性生物炭的(O+N/C)值增大,则其极性增强,与极性有机物的作用增强.乙草胺的苯环属于缺电子体系,可作为电子受体参与π-π电子供体受体作用.乙草胺中的氧可作为氢键的电子受体,与生物炭表面的含氧官能团和含氮组分形成氢键(王子莹等, 2016; 王菲等, 2016).
4 结论(Conclusions)1) 随着制备温度的升高,小麦和玉米秸秆生物炭的灰分含量、pH、CEC、比表面积增大,且小麦秸秆生物炭均大于玉米秸秆生物炭;生物炭芳香性增强、亲水性和极性减弱.改性生物炭样品的比表面积略微减小,CEC含量显著升高,极性和亲水性增强,官能团种类不变但数量增多,XRD谱图有明显的MgO衍射峰.
2) 改性生物炭XM-Mg(CH3COO)2的吸附性能最优,对氨氮、Pb(Ⅱ)和乙草胺3种吸附质的吸附量分别为6.57、16.81和9.79 mg·g-1.动力学研究表明:改性生物炭对氨氮、Pb(Ⅱ)和乙草胺的吸附均符合准二级吸附动力学;等温线研究表明:氨氮和乙草胺的的吸附过程均符合Langmuir模型,Pb(Ⅱ)的吸附过程符合Freundlich模型;热力学研究表明:低浓度时氨氮、Pb(Ⅱ)和乙草胺的吸附过程均可自发进行,升温有利于Pb(Ⅱ)和乙草胺的吸附.
3) 改性生物炭对氨氮、Pb(Ⅱ)的吸附以离子交换作用为主,对乙草胺的吸附以分配、氢键和π-π电子受体供体作用为主.
Ahmad M, Rajapaksha A U, Lim J E, et al. 2014. Biochar as a sorbent for contaminant management in soil and water:A review[J]. Chemosphere, 99(3): 19–33.
|
Cao X D, Harris W. 2010. Properties of dairy-manure-derived biochar pertinent to its potential use in remediation[J]. Bioresource Technology, 101(14): 5222–5228.
DOI:10.1016/j.biortech.2010.02.052
|
曹志宏, 黄艳丽, 郝晋珉. 2018. 中国作物秸秆资源利用潜力的多适宜性综合评价[J]. 环境科学研究, 2018, 31(1): 179–186.
|
Chen B L, Chen Z M, Lv S F. 2010. A novel magnetic biochar efficiently sorbs organic pollutants and phosphate[J]. Bioresource Technology, 102(2): 716–723.
|
程新伟. 2011. 土壤铅污染研究进展[J]. 地下水, 2011, 33(1): 5–68.
|
Chun Y, Sheng G Y, Chiou C T, et al. 2004. Compositions and sorptive properties of crop residue-derived chars[J]. Environmental Science & Technology, 38(17): 4649–4655.
|
董双快, 徐万里, 吴福飞, 等. 2016. 铁改性生物炭促进土壤砷形态转化抑制植物砷吸收[J]. 农业工程学报, 2016, 32(15): 204–212.
DOI:10.11975/j.issn.1002-6819.2016.15.028 |
丁绍兰, 屈振宇, 杨鹏. 2010. 改性核桃壳吸附废水中氨氮的机理研究[J]. 环境科学与技术, 2010(s2): 277–281.
|
Fang C, Zhang T, Li P, et al. 2014. Application of magnesium modified corn biochar for phosphorus removal and recovery from swine wastewater[J]. International Journal of Environmental Research & Public Health, 11(9): 9217–9237.
|
葛鑫, 戴其根, 霍中洋, 等. 2003. 农田氮素流失对环境的污染现状及防治对策[J]. 耕作与栽培, 2003, 26(6): 53–54.
|
何莉莉. 2013.生物炭对土壤微生态环境的影响及机理研究[D].北京: 中国林业科学研究院
http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y2405281 |
环境保护部, 国土资源部. 2014. 全国土壤污染状况调查公报[J]. 中国环保产业, 2014, 36(5): 10–11.
|
Ho Y S, McKay G. 1999. Pseudo-second order model for sorption processes[J]. Process Biochemistry, 34(5): 451–465.
DOI:10.1016/S0032-9592(98)00112-5
|
Keiluweit M, Nico P S, Johnson M G, et al. 2010. Dynamic molecular structure of plant biomass-derived black carbon (biochar)[J]. Environmental Science & Technology, 44(4): 1247–1253.
|
Lagergren S. 1898. About the theory of so-called adsorption of soluble substances[J]. Kungliga Svenska Vetensk. Handlingar, 24(4): 1–39.
|
Lee J, Kidder M, Evans B R, et al. 2010. Characterization of biochars produced from comstovers for soil amendment[J]. Environmental Science & Technology, 44(20): 7970–7974.
|
Lehmann J. 2007. A handful of carbon[J]. Nature, 447(7141): 143–144.
DOI:10.1038/447143a
|
罗小玲, 郭庆荣, 谢志宜, 等. 2014. 珠江三角洲地区典型农村土壤重金属污染现状分析[J]. 生态环境学报, 2014, 23(3): 485–489.
DOI:10.3969/j.issn.1674-5906.2014.03.018 |
李洋, 宋洋, 王芳, 等. 2015. 小麦秸秆生物炭对高氯代苯的吸附过程与机制研究[J]. 土壤学报, 2015, 52(5): 1096–1105.
|
马锋锋, 赵保卫. 2017a. 玉米芯生物炭吸附水中对硝基苯酚的特性[J]. 环境化学, 2017a, 36(4): 898–906.
|
马锋锋, 赵保卫, 刁静茹. 2017b. 小麦秸秆生物炭对水中Cd(Ⅱ)的吸附特性研究[J]. 中国环境科学, 2017b, 37(2): 551–559.
|
马锋锋, 赵保卫. 2017c. 不同热解温度制备的玉米芯生物炭对对硝基苯酚的吸附作用[J]. 环境科学, 2017c, 38(2): 837–844.
|
闵凡飞, 陈清如, 张明旭. 2006. 新鲜生物质热解气化半焦特性的XRD研究[J]. 中国矿业大学学报, 2006, 35(3): 336–340.
DOI:10.3321/j.issn:1000-1964.2006.03.010 |
孟繁健, 朱宇恩, 李华, 等. 2017. 改性生物炭负载nZVI对土壤Cr(Ⅵ)的修复差异研究[J]. 环境科学学报, 2017, 37(12): 4715–4723.
|
王菲, 孙红文. 2016. 生物炭对极性与非极性有机污染物的吸附机理[J]. 环境化学, 2016, 35(6): 1134–1141.
|
王连超. 2017. 污染土壤修复技术研究现状与趋势[J]. 环境与发展, 2017, 29(5): 68–69.
|
Wu W X, Yang M, Feng Q B, et al. 2012. Chemical characterization of rice straw-derived biochar for soil amendment[J]. Biomass & Bioenergy, 47(4): 268–276.
|
Wang X S, Tang Y P, Tao S R. 2008. Removal of Cr (Ⅵ) from aqueous solutions by the nonliving biomass of Alligator weed:kinetics and equilibrium[J]. Adsorption-journal of the International Adsorption Society, 14(6): 823–830.
DOI:10.1007/s10450-008-9145-6
|
王子莹, 邱梦怡, 杨妍, 等. 2016. 不同生物炭吸附乙草胺的特征及机理[J]. 农业环境科学学报, 2016, 35(1): 93–100.
|
薛俊杰. 2016.玉米、小麦秸秆原料、热解过程及固体产物特性NIRS快速分析研究[D].北京: 中国农业大学
http://cdmd.cnki.com.cn/Article/CDMD-10019-1016085334.htm |
谢淘. 2015.生物炭的特性分析及其在黄水资源化中的应用[D].北京: 清华大学
http://cdmd.cnki.com.cn/Article/CDMD-10003-1016712271.htm |
Yuan J H, Xu R K, Zhang H. 2011. The forms of alkalis in the biochar produced from crop residues at different temperatures[J]. Bioresource Technology, 102(3): 3488–3497.
DOI:10.1016/j.biortech.2010.11.018
|
郁强强. 2015.改性类水滑石和沼渣生物炭对磷/氨氮的吸附机理和应用研究[D].厦门: 厦门大学
http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y2926409 |
杨惟薇. 2014.生物炭对镉污染土壤的修复研究[D].南宁: 广西大学
http://kns.cnki.net/KCMS/detail/detail.aspx?filename=JSNY201506110&dbname=CJFD&dbcode=CJFQ |
赵玲, 滕应, 骆永明. 2017. 中国农田土壤农药污染现状和防控对策[J]. 土壤, 2017, 49(3): 417–427.
|
赵明静, 杜霞, 郭萌, 等. 2016. CaCl2改性生物炭的制备及其对Pb(Ⅱ)的吸附作用[J]. 环境污染与防治, 2016, 38(10): 84–88.
|
Zhang S Q, Yang X, Liu L, et al. 2018. Adsorption behavior of selective recognition functionalized biochar to Cd(Ⅱ) in wastewater[J]. Materials, 11(2): 299.
DOI:10.3390/ma11020299
|
朱银涛, 李业东, 王明玉, 等. 2018. 玉米秸秆碱化处理制备的生物炭吸附锌的特性研究[J]. 农业环境科学学报, 2018, 37(1): 179–185.
|