 2018, Vol. 38
2018, Vol. 38



2. 北京师范大学水科学研究院, 北京 100875;
3. 中国环境科学研究院, 环境基准与风险评估国家重点实验室, 流域水生态保护技术研究室, 北京 100012
2. College of Water Science, Beijing Normal University, Beijing 100875;
3. Laboratory of Riverine Ecological Conservation and Technology, State Key Laboratory of Environmental Criteria and Risk Assessment, Chinese Research Academy of Environmental Sciences, Beijing 100012
硅藻(Diatom)广泛存在河流、湖泊和水库中, 是天然水体中藻类群落的重要组成部分, 也是河流、湖泊和水库生态系统中的主要生产者(李丽娟等, 2015;刘麟菲等, 2014).硅藻对环境的变化有着极高的敏感性, 不同的河流底质类型会导致硅藻的群落结构有着不同的变化.河流底质是由非均匀的石块和泥沙颗粒组成, 属于一个多级分散的颗粒系统, 不同类型的底质为硅藻提供了附着基, 使硅藻着生在不同类型的河流底质中, 如石块、鹅卵石、砂砾、泥滩等底质类型(Wojtal et al., 2012).河流底质是影响硅藻群落结构的重要环境因素(Mike et al., 1993), 底质粒径的大小、异质性、松散程度和稳定性对硅藻群落结构的形成具有重要影响(Beisel et al., 2000;Jowett et al., 1990).
基于不同的河流底质类型并运用统计学方法可以建立河流底质指数(IOS)(闵文武等, 2016).根据计算出的河流底质指数, 可以推断出河流的底质类型, 数值越高说明河流底质复杂性越高, 底质较为松散, 其中, 松散型的底质多为较大的石块、鹅卵石等;数值越低说明河流底质复杂性单一, 河流底质较为密实, 密实型底质多为淤泥、砂砾等(Heino et al., 2000).近年来, 国内外学者研究了不同的底质类型对硅藻群落多样性的影响, 在国外的研究中, Kitner等(2003)与Marco等(2012)认为大石块底质类型的硅藻种群多样性要显著高于其它底质类型上的硅藻种群;Winter等(2000)则认为硅藻群落多样性受底质类型的影响不显著.在国内的研究中, 闵文武等(2016)通过硅藻对IOS指数的适应范围研究认为, 不同的硅藻种类所适应的河流底质类型并不相同;林金城等(2015)认为不同的河流基质上硅藻的种群有较大的差异性.基于国内外对于底质类型对硅藻群落多样性影响的探讨, 本研究旨在分析不同的河流底质是否是影响硅藻群落结构的主要因子.
浑河是辽河的支流, 位于大伙房水库上游区域, 承担着辽宁省中心城市的工农业生产及生活用水的水生态功能, 河流底质类型多数为较大的石块、鹅卵石等;而在浑河中下游地区, 底质类型多为沙和淤泥(殷旭旺等, 2011).由于浑河流域底质类型复杂, 因此, 本文以浑河为研究范例, 以河流硅藻群落为研究对象, 探讨不同的底质类型对硅藻群落结构及多样性的影响.
2 材料和方法(Materials and methods) 2.1 研究区域浑河水系(40°20′~41°00′N, 122°20′~125°20′E)位于我国辽宁省东部地区, 流域面积1.14×104 km2, 全长415 km.浑河发源于清源县, 与太子河交汇后形成大辽河, 最后经营口注入渤海, 其主要支流包括红河、英额河及苏子河等.为了研究浑河流域不同底质类型对硅藻群落结构的影响, 在浑河流域设定23个底质类型不同的点位进行调查(图 1).点位的选取原则主要是根据流速、水深、河流底质组成及其它环境因素, 在均有代表性的河段进行设定.各个点位的底质类型和采样点位水文特征等参数不同, 其环境条件见表 1.
 |
| 图 1 浑河流域采样点位示意图 Fig. 1 Locations of sampling sites in the Hunhe River |
| 表 1 采样点底质类型及水文参数 Table 1 Sediment type and hydrological parameter of sampling point |
根据附着硅藻栖地的复杂性, 在野外采集硅藻样品时应该采集定量和定性样品各一份.在采样点河流上下游100 m范围内, 根据河流生境不同(流速、水深和透明度等), 挑选9个不同生境的石块, 在每一个石块上用硬毛刷分别刮取面积为15.2 cm2(直径4.4 cm的圆形塑胶环)的着生藻类, 使用自来水将其冲刷至不锈钢托盘中, 将定量混合的硅藻倒入广口塑料瓶中, 加入5%(体积分数)的甲醛固定, 定性的样品是将刚采集过不同生境上的剩余硅藻全部刷至不锈钢托盘中, 倒入另一广口塑料瓶中, 加入5%的甲醛固定.若底质没有石块, 则根据泥沙底质分3个不同的区域, 刮取表面上的泥沙(约2 cm厚)放进广口塑料瓶中, 加入甲醛(5%)固定, 刮取的样品面积为36.32 cm2(底面直径6.8 cm的钟形不锈钢盖), 同样需要采集1份定量样品和1份定性样品.采集到的着生藻类样品先浓缩处理, 定性的样品要进行酸化处理, 并且需要制作着生藻类的封片, 最后于1000×显微镜下计数1000个着生藻类细胞.定量样品直接定量到100 mL, 于400×显微镜下直接进行细胞计数, 每片计数100个视野.种类鉴定依据相关文献(朱蕙忠等, 2000;胡鸿钧等, 2006).
2.3 底质指数及水环境理化指标的测定针对河流底质的采集, 采用分样筛和量杯野外现场测定不同粒径底质的体积.底质标准按照不同粒径分为:漂石(>256 mm)、大卵石(128~256 mm)、小卵石(64~128 mm)、大圆石(32~64 mm)、小圆石(16~32 mm)、粗砾(8~16 mm)、细砾(4~8 mm)和砂(<4 mm)(Winter et al., 2000).底质指数(IOS)根据Allan等(2007)公式计算:IOS=0.08%V漂石+0.07%V大卵石+0.06%V小卵石+0.05%V大圆石+0.04%V小圆石+0.03%V粗砾 +0.02%V细砾+0.01%V砂.
浑河各采样点位的电导率(Cond)、总溶解固体(TDS)和溶解氧(DO)均用水质分析仪(YSI Pro 2000)现场测定;水深和流速(CS)使用流速仪现场测定;另外采集一份2 L水样, 在低温条件下(4 ℃)送回实验室, 根据标准方法(国家环境保护总局, 2002), 测定水化学指标, 包括:氨氮(NH4+-N)、总氮(TN)、总磷(TP)和高锰酸盐指数(CODMn)等.
2.4 数据分析应用多响应置换过程(Multi-Response permutation procedures, MRPP)分析苏子河、红河、英额河、浑河中游及浑河下游的硅藻群落结构的空间异质性.计算浑河流域各个采样点位硅藻的物种丰富度、香农多样性指数(H)(Shannon et al., 1963)及均匀度指数(J)(Pielou et al., 1963), 具体公式如下:
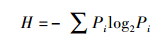
|
(1) |

|
(2) |
式中, Pi为第i个物种的个体数与所有种类个体数的比值;Hmax= log2S, S为样品中总物种数.对浑河流域底质指数在不同河段下进行单因素方差分析(one-way ANOVA)和独立样本T检验, p值小于0.05, 则数据差异性显著.对硅藻物种密度进行除趋势对应分析(Detrended Correspondence Analysis, DCA), 可以得出物种的单峰影响值梯度SD, 若SD>2, 则进行典范对应分析(Canonical Correspondence Analysis, CCA), 若SD<2, 则进行冗余度分析(Redundancy Correspondence Analysis, RDA).同时对数据进行9999次蒙特卡洛置换检验(Monte Carlo Test, MCT), 来判断显著影响浑河流域硅藻群落空间分布特征的环境因子(p<0.05).通过柯尔莫哥诺夫-斯米尔诺夫分布检验(Kolmogorov-Smirnov Test, K-S), 分析香农多样性指数、均匀度指数及物种丰富度是否符合正态分布, 如符合正态分布, 则相关分析判断香农多样性指数、均匀度指数、物种丰富度与底质指数(IOS)是否具有相同的趋势变化.
采用PC-ORD 5.0进行MRPP分析;箱型图、线形回归拟合分析均在Origin 7.5上完成;采用Canoco 4.5进行除趋势对应分析和典范对应分析;采用SPSS 19.0进行单因素方差分析;除pH外, 所有的水环境数据和硅藻群落结构数据进行数据转化lg(x+1).
3 结果(Results) 3.1 硅藻群落分布特征在本次采样调查中, 浑河流域共鉴定出硅藻84种, 其中, 常见硅藻种类有:谷皮菱形藻(Nitzschia palea)、偏肿桥弯藻(Cymbella ventricosa)、膨大桥弯藻(C. turgida)、弧形峨眉藻(Ceratoneis arcus)、线形菱形藻(N. linearis)、简单舟形藻(Navicula simplex)、小片菱形藻(N. frustulum)、梅尼小环藻(Cyclotella meneghiniana)等, 分别占总个体数的22.4%、9.8%、8.8%、6.8%、5.2%、3.8%、3.8%、3.2%.
MRPP结果显示, 苏子河、红河和英额河、浑河中游及浑河下游水系之间硅藻群落空间异质差异显著(p<0.0001).表 2结果显示, 不同河段下群落结构组成的差异比较明显.苏子河采样点位共鉴定出43种硅藻, 其中以膨大桥弯藻、弧形峨眉藻、偏肿桥弯藻和平卧桥弯藻等物种居多;红河和英额河采样点位共鉴定出52种硅藻, 其中以偏肿桥弯藻、膨大桥弯藻、谷皮菱形藻、简单舟形藻和泉生菱形藻等物种居多;浑河中游采样点位共鉴定出55种硅藻, 其中以弧形峨眉藻、偏肿桥弯藻、膨大桥弯藻和小片菱形藻等物种居多;浑河下游采样点位共鉴定出硅藻35种, 其中谷皮菱形藻、线形菱形藻、梅尼小环藻和链形小环藻等物种居多.
| 表 2 浑河流域不同河段中硅藻密度 Table 2 Diatom density of different river in Hunhe River |
对于浑河流域硅藻群落结构, 香农多样性指数、均匀度指数与物种丰富度的平均值分别为2.57、0.69和15.图 2结果显示, 硅藻香农多样性指数与均匀度指数从高到低依次为:红河和英额河>苏子河>浑河中游>浑河下游; 而物种丰富度从高到低依次为:红河和英额河>苏子河>浑河下游>浑河中游.在不同河段硅藻香农多样性指数中, 苏子河与浑河中游、苏子河与浑河下游、红河和英额河与浑河中游、红河和英额河与浑河下游、浑河中游与浑河下游呈显著相关性(p<0.05);在不同河段硅藻均匀度指数中, 苏子河与浑河中游、苏子河与浑河下游、红河和英额河与浑河下游呈显著相关性(p<0.05);在不同河段硅藻的物种丰富度中, 苏子河与浑河中游、苏子河与浑河下游、红河和英额河与浑河中游、红河和英额河与浑河下游、浑河中游与浑河下游均呈显著相关性(p<0.05).
 |
| 图 2 不同河段中硅藻群落结构的变化 (A1:苏子河;A2:红河和英额河;A3:浑河中游;A4:浑河下游) Fig. 2 Community structure of diatom in different rivers |
单因素方差结果显示(表 3), 除苏子河、红河和英额河之间底质指数差异不显著(p>0.05), 其余水系之间底质指数差异均显著(p<0.05).底质指数最高值出现在苏子河, 最低值出现在浑河下游, 苏子河、红河和英额河底质指数较高, 底质类型较浑河中游和浑河下游相对复杂, 浑河中游和浑河下游底质类型较为单一.
| 表 3 浑河流域4种不同河段中底质指数的比较 Table 3 Comparison of IOS of four rivers in the Hun River |
对浑河流域23个采样点位进行除趋势对应分析(DCA), 其物种的单峰影响值梯度为4.108(SD=4.108, SD>2), 表明浑河流域硅藻群落结构对生态梯度的响应是非线性的, 因此, 采用典范对应性分析(CCA)来分析环境因子对硅藻群落结构的影响.CCA结果显示(图 3), IOS指数(p=0.0047, F=1.675)、电导率(p=0.0008, F=1.869)、总氮(p=0.0037, F=1.694)和总磷(p=0.0283, F=1.520)对硅藻群落结构的影响要高于溶解氧(p=0.7347, F=0.866).
 |
| 图 3 浑河流域水环境理化因子与硅藻群落的典范对应分析 Fig. 3 Canonical correspondence analysis (CCA) of species-environmental relationships in the Hunhe River |
通过K-S分布检验, 得出香农多样性指数、均匀度指数和物种丰富度均呈正态分布, 因此, 对硅藻群落结构与河流底质指数(IOS)之间进行相关分析(图 4).结果显示, 香农多样性指数、均匀度指数和物种丰富度均与河流底质指数(IOS)呈正相关, 随着IOS指数数值的增加, 香农多样性指数、均匀度指数和物种丰富度数值也随之增加, 且香农多样性指数与IOS指数之间的相关系数最高(r=0.675;p<0.001).
 |
| 图 4 硅藻群落结构与IOS指数的相关分析 Fig. 4 Correlation analysis of diatom community structure with IOS |
浑河流域各个点位硅藻物种所占的百分比(表 4)显示, 苏子河水系底质指数范围为2.300~8.055, 且各个点位的优势物种多为偏肿桥弯藻C. Ventricosa和平卧桥弯藻C. Prostrata;红河和英额河流域底质指数范围为2.770~6.755, 且各个点位的优势物种多为偏肿桥弯藻C. Ventricosa和谷皮菱形藻N. Palea;浑河中游底质指数范围为0.40~1.67, 各个点位的优势物种多为谷皮菱形藻N. Palea和弧形峨眉藻C. arcus;浑河下游底质指数范围为0.2~0.6, 各个点位的优势物种多为梅尼小环藻C. meneghiniana和谷皮菱形藻N. Palea.
| 表 4 各个点位主要物种所占百分比 Table 4 Percentage of major species at each site |
殷旭旺等(2011)在2010年对浑河水系进行了采样调查, 鉴定出硅藻共134种.而本研究在浑河流域共采集到硅藻84种, 远远小于殷旭旺等在浑河流域采样调查的硅藻物种, 同时也小于太子河(李丽娟等, 2015;闵文武等, 2016)、冈曲河(Wu et al., 2009)和香溪河(Tang et al., 2004)等其他流域.研究表明, 相比于其他河流环境因子, 河流底质类型对硅藻群落结构及多样性有着更大的影响, 稳定的河流底质可以为硅藻提供良好的生境, 但采矿、挖沙、水体污染等会导致河道底质泥沙堆积, 使硅藻丧失其原有的生境, 导致硅藻的物种数量及密度明显下降(殷旭旺等, 2012).在本研究中, 浑河中游有很多采矿和挖沙等现象, 而浑河下游由于人类活动频繁, 水体污染也较为严重, 底质类型多为淤泥, 硅藻群落多样性降低甚至丧失.相反, 苏子河、红河和英额河人为干扰较少, 森林覆盖率高, 底质类型多为大石块和鹅卵石等, 为硅藻提供了稳定的基质, 所以硅藻的多样性也相对较高.
本研究发现, 苏子河、红河和英额河底质类型较为复杂, 多为大石块和鹅卵石, 底质较为稳定, 且优势物种为偏肿桥弯藻C. Ventricosa和平卧桥弯藻C. Prostrata;浑河中游和浑河下游底质类型比较单一, 多为泥沙和淤泥, 底质不稳定, 且优势物种为谷皮菱形藻N. Palea和梅尼小环藻C. Meneghiniana, 并且在H12和H19采样站位, 谷皮菱形藻分别占该点位所有物种的78.8%和75.8%.这可能是由于硅藻对不同的河流底质类型具有不同的适应性, 并且其自身的形态和能动性具有一定的关系(Stenger et al., 2013).例如, 菱形藻属属于运动型硅藻(殷旭旺等, 2012), 其运动能力较强, 既可以附着在泥沙底质的表面, 也可附着在石块、鹅卵石及砂砾的河流底质类型中, 所以在浑河流域中广泛分布, 而桥弯藻属不具备运动能力(殷旭旺等, 2012), 其附着基质多为较大的石块, 所以广泛分布于苏子河、红河和英额河.
单因素方差分析结果显示, 除苏子河、红河和英额河之间底质指数差异不显著, 其余水系之间底质指数差异均显著.这种造成不同河段底质指数之间差异的原因可能是:苏子河、红河和英额河受到人为的干扰较小, 森林覆盖率较大, 底质类型多为大型的石块和鹅卵石等, 仅有少数的点位底质类型为砂砾, 水中溶解氧较高, 硅藻多样性较高.浑河中游由于个别点位出现采沙和开矿现象, 导致河流基质被细沙覆盖, 使一些运动能力弱的桥弯藻属无法附着在基质上, 仅有运动能力较强的菱形藻属和舟形藻属在浑河中游广泛分布(Stenger et al., 2013).浑河下游由于水体的污染相对严重, 底质多为淤泥等富营养化沉积物, 因此, 指示水体贫营养状态的藻类, 如膨大桥弯藻、窄异极藻(Kovacs et al., 2007)等物种在浑河下游没有被发现, 而指示富营养化的谷皮菱形藻和指示多污染带的梅尼小环藻(Dong et al., 2006)则大量出现, 因此, 浑河下游硅藻多样性比较单一.
根据CCA结果显示, IOS指数、电导率、总氮和总磷是显著影响硅藻群落结构的驱动因子.而对于不同的流域, 其影响硅藻群落结构的驱动因子也并不相同, 如太子河流域中, IOS指数、总溶解固体和高锰酸盐指数为显著影响硅藻群落结构的驱动因子(闵文武等, 2016), 在桂江流域中, 氨氮、硝酸碳氮和总氮是显著影响硅藻群落结构的主要因子(邓培雁等, 2012).本研究中IOS指数与硅藻香农多样性指数、均匀度指数和物种丰富度呈现显著正相关性.有研究表明(Stevenson et al., 1996;刘霞等, 2012), 硅藻在进行光合作用时, 需要氮、磷元素作为营养元素, Leland等(2000)研究认为, 氨氮和总磷是影响着生藻类群落结构的主要环境因子, 本研究结果与之一致.在浑河流域中, 苏子河、红河和英额河与浑河中游、浑河下游不同的氮、磷浓度可能是影响硅藻群落多样性显著差异的主要原因.电导率是水中总溶解离子量的参数, 它通常会伴随着人类活动的频繁而升高, 特别是在城镇用地和耕地下的土地利用类型, 参数数值显著升高(Walker et al., 2006).本研究显示, 苏子河、红河和英额河流域人类活动较少, 森林覆盖率较大, 电导率较低, 硅藻多样性较高;在浑河中游及浑河流域下游由于人类活动频繁, 出现耕地和城镇用地, 森林覆盖率低, 导致电导率升高, 硅藻多样性较低甚至丧失.
Kumar等(2009)认为, 稳定的河流底质可以提高物种多样性, 而不稳定的河流底质对水生生物群落结构有着不利的影响(Beisel et al., 1998;Cummins et al., 1979).Biggs等(2002)认为底质类型为石块、鹅卵石等基质可为着生藻类提供稳定的附着基, 而砂砾和泥沙等基质不利于着生藻类的附着.在本研究中, 底质指数与硅藻香农多样性指数、物种丰富度和均匀度指数之间呈现正相关性, 说明底质若为石块鹅卵石等类型, 其硅藻物种的多样性也就越高, 这与Biggs等(2002)的研究相一致.浑河流域上游的苏子河、红河和英额河及浑河的底质类型较为复杂, 底质多为石块鹅卵石, 具有较高的稳定性, 并且较大粒径的石块可以为硅藻提供很好的附着基质, 不会随着水流冲到下游, 所以着生硅藻的群落结构比较稳定.浑河下游的底质类型多为泥沙, 流速对其影响较大, 泥沙随着水流流动, 导致附着在泥沙表面上的硅藻被一起带走, 从而破坏硅藻的群落结构, 导致多样性的丧失.
5 结论(Conclusions)1) 在本次采样调查中, 浑河流域共鉴定出硅藻84种, 其中, 常见硅藻种类有:谷皮菱形藻、偏肿桥弯藻、膨大桥弯藻、弧形峨眉藻、线形菱形藻、简单舟形藻、小片菱形藻、梅尼小环藻等, 分别占总个体数的22.4%、9.8%、8.8%、6.8%、5.2%、3.8%、3.8%、3.2%.
2) 浑河流域硅藻群落结构, 香农多样性指数、均匀度指数与物种丰富度的平均值分别为2.57、0.69和15.硅藻香农多样性指数与均匀度指数从高到低依次为:红河和英额河>苏子河>浑河中游>浑河下游.而物种丰富度从高到低依次为:红河和英额河>苏子河>浑河下游>浑河中游.
3) IOS指数、电导率、总氮和总磷是显著影响硅藻群落结构的环境因子.香农多样性指数、均匀度指数和物种丰富度均与河流底质指数呈现出正相关关系, 并具有相同的变化趋势, 随着IOS指数数值的增加, 香农多样性指数、均匀度指数和物种丰富度数值也随之增加.
Allan J D, Castillo M M. 2007. Stream Ecology:Structure and Function of Running Waters(2nd ed)[M]. Netherlands: Springer.
|
Beisel J N, Usseglio-Polatera P, Moreteau J C. 2000. The spatial heterogeneity of a river bottom:a key factor determining macroinvertebrate communities[J]. Hydrobiologia, 422/423: 163–171.
DOI:10.1023/A:1017094606335
|
Beisel J N, Usseglio-Polatera P, Thomas S, et al. 1998. Stream community structure in relation to spatial variation:the influence of mesohabitat characteristics[J]. Hydrobiologia, 389(1): 73–88.
|
Biggs B J F, Smith R A. 2002. Taxonomic richness of stream benthic algae:effects of fflood disturbance and nutrients[J]. Limnology and Oceanography, 47: 1175–1186.
DOI:10.4319/lo.2002.47.4.1175
|
Cummins K M. 1979. The Natural Stream Ecosystem//Ward J V, Stanford J A eds. The Ecology of Regulated Stream[M]. New York: Plenum Press. 7-24
|
邓培雁, 雷远达, 刘威, 等. 2012. 桂江流域附生硅藻群落特征及影响因素[J]. 生态学报, 2012, 32(7): 2196–2203.
|
Dong X H, Yang X D, Wang R. 2006. Diatom indicative species of eutrophication of the lakes in the middle and lower reach regions of Yangtze River[J]. China Environmental Science, 26(5): 570–574.
|
国家环境保护总局. 2002. 水和废水监测分析方法(第4版)[M]. 北京: 中国环境科学出版社.
|
Heino J. 2000. Lentic macroinvertebrate assemblage structure along gradients in spatial heterogeneity, habitat size and water chemistry[J]. Hydrobiologia, 418: 229–242.
DOI:10.1023/A:1003969217686
|
胡鸿钧, 魏印心. 2006. 中国淡水藻类:系统分类及生态[M]. 北京: 科学出版社.
|
Jowett I G, Richardson J. 1990. Microhabitat preference of benthic invertebrates in a New Zealand river and the development of instream flow-habitat models for Deleatidum spp[J]. New Zeal Journal of Marine Freshwater Research, 24: 19–30.
DOI:10.1080/00288330.1990.9516399
|
Kitner M, Poulícková A. 2003. Littoral diatoms as indicators for the eutrophication of shallow lakes[J]. Hydrobiologia, 506/509(1/3): 519–524.
|
Kovacs C S, Buczko K, Hajnal E. 2007. Epiphytic, littoral diatoms as bioindicators of shallow lake trophic status:Trophic diatom index for lakes (TDIL) developed in Hungary[J]. Hydrobiologia, 589: 141–154.
DOI:10.1007/s10750-007-0729-z
|
Kumar S, Spaulding S A, Stohlgren T J, et al. 2009. Potential habitat distribution for the freshwater diatom Didymosphenia geminate in the continental US[J]. Frontiers Ecology Environmental, 8(7): 415–420.
|
Leland H V, Porter S D. 2000. Distribution of benthic algae in the upper Illinois River basin in relation to geology and land use[J]. Freshwater Biology, 44: 279–301.
DOI:10.1046/j.1365-2427.2000.00536.x
|
李丽娟, 金文, 王博涵, 等. 2015. 太子河河岸土地利用类型与硅藻群落结构的关系[J]. 环境科学研究, 2015, 28(11): 1662–1669.
|
林金城, 蔡德所, 姚文婷, 等. 2015. 硅藻群落结构的差异:比较样本采集过程与空间梯度[J]. 湖泊科学, 2015, 27(3): 407–411.
DOI:10.18307/2015.0306 |
刘麟菲, 谭冰冰, 殷旭旺, 等. 2014. 在种与属两个级别评价太子河硅藻群落与环境因子的关系[J]. 生态学报, 2014, 34(22): 6613–6621.
|
刘霞, 陆晓华, 陈宇炜. 2012. 太湖浮游硅藻时空演化与环境因子的关系[J]. 环境科学学报, 2012, 32(4): 821–827.
|
Marco C, Nicola A, Ermanno B, et al. 2012. Diatoms in spring of the Alps:spring types, environmental determinants, and substratum[J]. Freshwater Science, 31(2): 499–524.
DOI:10.1899/11-065.1
|
Scarsbrook Mike R., Townsend Colin R.. 1993. Stream community structure in relation to spatial and temporal variation:a habitat templet study of two contrasting New Zealand streams[J]. Freshwater Biology: 29, 395–410.
|
闵文武, 韩洁, 王培培, 等. 2016. 太子河流域硅藻群落与驱动因子的定量关系[J]. 环境科学研究, 2016, 29(5): 672–679.
|
Pielou E C. 1996. Species-diversity and pattern-diversity in the study of ecological succession[J]. Journal of Theoretical Biology, 10: 370–380.
|
Shannon C E, Weaver W. 1963. The Mathematical Theory of Communication[M]. Urbana: University of Illinois Press.
|
Stenger K C, Lenggel E, Crossetti L O, et al. 2013. Diatom ecological guilds as indicators of temporally changing stressors and disturbances in the small Torna-stream, Hungary[J]. Ecology Indicators, 24: 138–147.
DOI:10.1016/j.ecolind.2012.06.003
|
Stevenson R J, Bothwell M L, Lowe R L. 1996. Algal Ecology:Freshwater Benthic Ecosystems[M]. San Diego, USA: Academic Press.
|
Tang T, Qu X D, Li D F, et al. 2004. Benthic algae of the Xiangxi River, China[J]. Journal of Freshwater Ecology, 19(4): 597–604.
DOI:10.1080/02705060.2004.9664740
|
Walker C E, Pan Y D. 2006. Using diatom assemblages to assess urban stream conditions[J]. Hydrobiologia, 561(1): 179–189.
DOI:10.1007/s10750-005-1613-3
|
Winter J G, Duthie H C. 2000. Stream epilithic, epipelic and epiphytic diatoms:habitat fidelity and use in biomonitoring. Aquatic[J]. Ecology, 34: 345–353.
|
Wu N C, Tang T, Zhou S C, et al. 2009. Changes in benthic algal communities following construction of a run-ofriver dam[J]. Journal of the North American Benthological Society, 28(1): 69–79.
DOI:10.1899/08-047.1
|
殷旭旺, 渠晓东, 李庆南, 等. 2012. 基于着生藻类的太子河流域水生态系统健康评价[J]. 生态学报, 2012, 32(6): 1677–1691.
|
殷旭旺, 张远, 渠晓东, 等. 2011. 浑河水系着生藻类的群落结构与生物完整性[J]. 应用生态学报, 2011, 22(10): 2732–2740.
|
朱蕙忠, 陈嘉佑. 2000. 中国西藏硅藻[M]. 北京: 科学出版社.
|
Wojtal A Z, Sobczyk Ł. 2012. The influence of substrates and physicochemical factors on the composition of diatom assemblages in karst springs and their applicability in water-quality assessment[J]. Hydrobiologia, 695(1): 97–108.
DOI:10.1007/s10750-012-1203-0
|