 2018, Vol. 38
2018, Vol. 38



2. 湿润亚热带生态地理过程教育部重点实验室, 福州 350007
2. Key Laboratory of Humid Sub-tropical Eco-geographical Process of Ministry of Education, Fuzhou 350007
碳、氮、磷是土壤中主要的元素组成, 也是维持生态系统生产力的关键因子(Elser et al., 2007; Cleveland et al., 2007; Yang et al., 2010).近年来, 关于土壤碳、氮、磷的研究在关注其含量及周转过程的同时, 也开始逐步探讨不同元素之间的平衡关系(Elser et al., 2000).生态化学计量学主要是基于元素比率视角探讨生态系统能量和多元素平衡之间的关系(Elser et al., 2000).元素生态化学计量学研究已经成为当前研究的热点(Allen et al., 2009).
从国内外研究进展来看, 关于生态化学计量学的研究国外案例相对较多且较为深入(Elser et al., 2000; Tessier et al., 2003).虽然近年来国内学者也从不同角度研究了生态化学计量学特征的空间变异性及其指示作用等, 但相关研究仍显不足(Han et al., 2005; 王绍强等, 2008; 王维奇等, 2010).尤其是位于海陆交互作用的河口湿地元素生态化学计量学的研究更显不足, 实际上湿地土壤生态化学计量学特征在指示植物生长的限制性养分、生态系统服务功能及对外界环境的响应等方面均具有重要意义(Elser et al., 2007; 王维奇等, 2010).特别是受到海陆交互作用、周期性潮汐影响及土壤动植物干扰下的河口湿地, 其元素生态化学计量学特征更具复杂性与空间变异性, 该生态系统土壤碳、氮、磷耦合关系及响应机制应该受到更多关注, 深入开展相关研究对于厘清河口湿地的生态化学计量学特征与完善生态化学计量学理论均具有重要的意义.
底栖动物是河口湿地生态系统的次级生产者, 也是湿地生态系统的重要组成部分(Stahl et al., 2014; Vermeiren et al., 2015; Hu et al., 2016), 深刻影响着河口湿地生态系统物质循环和能量流动.底栖动物通过生物扰动(潜穴、爬行、觅食和避敌等)、排泄等生理活动和“生物灌溉”强烈改变着湿地生物地球化学循环过程, 特别是土壤碳、氮元素的循环(刘敏等, 2005; 胡知渊等, 2009), 但对磷元素的影响如何?目前仍鲜见报道.综上所述, 底栖动物对河口湿地土壤理化性质与养分的影响是十分复杂的.湿地底栖动物的活动对土壤碳、氮、磷含量及其生态化学计量学特征的影响在时间和空间上的表现如何, 值得开展研究.
螃蟹是河口湿地最为重要的底栖动物, 因此, 开展螃蟹对河口湿地土壤碳、氮、磷元素含量及其生态化学计量学特征的影响研究, 可为科学估算河口湿地碳、氮、磷元素含量及其空间变异性提供科学依据, 并为后续开展湿地底栖生物生态影响的相关研究积累经验, 同时也对湿地生态化学计量学的发展及其在湿地生态系统的应用具有重要意义.
2 研究区与研究方法(Study area and methods) 2.1 研究区概况该区属亚热带海洋性季风气候, 气候暖热湿润, 年均温为19.85 ℃, 平均年降雨量为1380 mm左右, 降水集中在3—9月, 表现为典型双峰变化, 6月为梅雨期, 8月为台风期(刘剑秋等, 2006).湿地土壤为盐渍化及脱盐化交替状态, pH呈偏酸性到中性, 粒径以粉砂为主, 砂砾含量较少.主要群落类型包括:秋茄群落(Kandelia candel)、木麻黄群落(Casuarina equisetifolia)、芦苇群落(Phragmites australis L.)、短叶茳芏群落(Cyperus malaccensis)、藨草群落(Scirpustriqueter)、互花米草群落(Spartina alterniflora)等.
本研究以闽江河口互花米草湿地为研究区, 选取相手蟹为研究对象, 相手蟹属于杂食种类, 除了取食植物的种子、茎叶外, 也可捕食各种小型的甲壳动物、软体动物和多毛类环节动物等(刘剑秋等, 2006).采样点的螃蟹洞穴以平洞和烟囱洞为主, 具有高潮滩的螃蟹洞直径大、但数量少, 中潮滩的螃蟹洞直径小、数量多的特征.
2.2 土壤样品采集在互花米草湿地内分别设置高潮滩有螃蟹影响的样地(简称为GP)、高潮滩无螃蟹影响的样地(简称为GD)、中潮滩有螃蟹影响的样地(简称为ZP)和中潮滩无螃蟹影响的样地(简称为ZD)4种处理, 每种处理设置3个重复.对于对照组, 为隔离螃蟹的影响, 分别用铁丝网(防锈)和PPR管进行围护(地上部分70 cm, 地下部分30 cm), 防止蟹类从地下掘穴进入, 样方大小为1 m×1 m.95%以上的大型底栖动物个体都分布在土壤表层10 cm深度范围内, 表层10 cm深处的螃蟹洞口直径较大, 向深处逐渐变小, 且螃蟹洞并不是一直垂直向下, 而是在10 cm左右就出现侧向延伸.考虑到洞穴特征和采样可行性, 故本研究仅采集土壤表层0~10 cm土壤样品, 分别于2015年冬季(1月)、春季(4月)、夏季(6月)和秋季(9月)采集, 每个样地利用土壤采样器多点采集, 迅速混合(降低土壤异质性及螃蟹洞的干扰)(王霞等, 2008).样品采集后, 立即放入自封袋, 并带回实验室, 去除杂质, 经自然风干后, 过2 mm筛, 之后再取部分样品过100目(0.149 mm)孔筛, 保存, 待测, 用于测量土壤全碳(TC)、全氮(TN)、全磷(TP)等指标(曾从盛等, 2008).
土壤全碳、全氮采用土壤碳氮元素分析仪测定(Vario EL, 德国, Zhou et al., 2007), 土壤全磷采用钼锑抗比色法(UV-2450)进行测定, 土壤含水量用烘干法测定, 土壤pH在土壤:水=1:5条件下用便携式IQ 150pH计测定, 土壤电导率(EC)及土壤温度采用2265FS电导仪原位测定, 土壤容重用环刀法进行测定, 具体测定方法参考文献(鲁如坤, 2000).
2.3 土壤碳、氮、磷储量计算方法土壤表层剖面单位面积一定深度碳、氮、磷储量(TC, t·km-2)用下式计算:
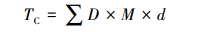
|
(1) |
式中, D为土壤表层容重(g·cm-3), M为土壤表层碳、氮、磷含量(g·kg-1), d为土层深度(cm)
2.4 数据处理运用Excel 2010、Origin、SPSS 20.0统计分析软件对测定数据进行整理.原始数据的平均值及标准误差采用Excel 2010计算, 对于不同处理样地的土壤全碳、全氮、全磷含量及其生态化学计量学特征等测定指标, 采用Origin 8.0软件做图, 土壤C/N、C/P、N/P比采用质量比, 螃蟹及对照土壤碳、氮、磷含量、储量及C/N、C/P、N/P比的差异性检验采用SPSS 20.0的重复测量方差分析方法, 环境因子与碳、氮、磷含量及C/N、C/P、N/P比, 以及C/N、C/P、N/P比与土壤碳、氮、磷储量的相关性分析采用SPSS 20.0的Pearson相关性分析.
3 结果与分析(Results and analysis) 3.1 螃蟹对土壤碳、氮、磷含量的影响螃蟹影响下土壤碳、氮、磷含量的季节变化见图 1, GP(高潮滩有螃蟹)组土壤全碳(TC)含量在11.42~15.17 g·kg-1之间, 平均值为(13.32±0.66) g·kg-1, 而GD(高潮滩无螃蟹)组土壤TC含量在11.33~15.39 g·kg-1之间, 平均值为(13.25±0.53) g·kg-1, GP组土壤TC含量与GD组差异不显著(p>0.05);而ZP(中潮滩有螃蟹)组土壤TC含量在11.80~15.48 g·kg-1之间, 均值为(13.85±0.61) g·kg-1, ZD(中潮滩无螃蟹)组土壤TC含量在11.66~13.52 g·kg-1之间, 均值为(13.10±0.43) g·kg-1, ZP组土壤TC含量显著高于ZD组(p<0.05), 说明在中潮滩有螃蟹活动下土壤碳含量相对较高, 变化较为明显.
 |
| 图 1 螃蟹对土壤全碳(TC)、土壤全氮(TN)和土壤全磷(TP)含量的影响 (图中不同大写字母表示同一季节不同处理间差异显著(p<0.05), 不同小写字母表示同一处理不同季节间差异显著(p<0.05), 下同) Fig. 1 Effect of crabs on soil TC, TN and TP concentration |
GP组土壤TN含量在1.28~1.75 g·kg-1之间, 平均值为(1.51±0.05) g·kg-1, GD组土壤TN含量在1.33~1.81 g·kg-1之间, 平均值为(1.52±0.04) g·kg-1, GP组土壤TN含量与GD组差异不显著(p>0.05).ZP组土壤TN含量在1.45~1.84 g·kg-1之间, 平均值为(1.64±0.08) g·kg-1, ZD组土壤TN含量在1.30~1.68 g·kg-1之间, 平均值为(1.52±0.08) g·kg-1, ZP组土壤TN含量显著高于ZD组(p<0.01).说明中潮滩有螃蟹活动下土壤氮含量相对较高, 变化较为明显.
GP组土壤TP含量在0.63~0.83 g·kg-1之间, 平均值为(0.70±0.05) g·kg-1, 而GD组土壤TP含量在0.66~0.87 g·kg-1之间, 平均值为(0.75±0.04) g·kg-1, GP组土壤TP含量与GD组差异不显著(p>0.05);而ZP组土壤TP含量在0.67~0.88 g·kg-1之间, 平均值为(0.77±0.07) g·kg-1, ZD组土壤TP含量在0.68~0.89 g·kg-1之间, 平均值为(0.79±0.04) g·kg-1, ZP组土壤TP含量与ZD组差异不显著(p>0.05).如表 1所示, 在高潮滩和中潮滩, 不管是螃蟹组还是对照组, 土壤TP都与季节存在极显著关系(p<0.05), 也就是在环境因子作用下, 土壤磷含量的变化较为明显.此外, 冬、春两季土壤碳、氮含量中, 高潮滩处理下的含量比中潮滩处理下的高(p<0.05), 而夏、秋两季的差异不显著(p>0.05).
| 表 1 土壤碳、氮和磷含量重复测量方差分析 Table 1 Repeated measure analysis of variance of soil TC, TN and TP concentration |
通过对3种元素的线性回归分析发现, 碳、氮、磷之间存在显著的线性拟合关系(p<0.01)(图 2), 从斜率上看, 碳和氮、氮与磷元素之间呈现良好的线性拟合的关系, 而碳与磷元素之间线性拟合的程度相对较低, 其中, 碳和氮几乎是同步变化的, 整体而言, 碳与氮元素变化较磷快.
 |
| 图 2 螃蟹对土壤碳、氮、磷线性拟合关系的影响 Fig. 2 Effect of crabs on soil C, N and P linear fit relationship |
由图 3可知,GP组土壤C/N比在8.29~9.30之间, 平均值为8.80±0.26, 变异系数为2.93%;C/P比在18.26~21.58之间, 平均值为19.38±1.34, 变异系数为6.93%;N/P比在2.07~2.32之间, 平均值为2.17±0.16, 变异系数为7.39%;GP组土壤C/N比相对稳定, 而C/P比和N/P比变化较大.GD组土壤C/N比在8.27~9.19之间, 平均值为8.70±0.28, 变异系数为3.23%;C/P比在16.46~19.00之间, 平均值为17.78±0.42, 变异系数为2.35%;N/P比在1.99~2.10之间, 平均值为2.04±0.13, 变异系数为6.41%;GD组土壤C/N比和C/P比相对稳定, 而N/P比变化较大.在高潮滩中, 螃蟹作用下3种比值均大于对照组, 如表 2所示, 土壤C/N比与不同季节呈显著关系(p<0.01), 土壤C/P比分别与不同季节和螃蟹作用呈现较为显著关系(p<0.05), 可见有螃蟹活动对土壤养分比的影响大.
 |
| 图 3 螃蟹对土壤C/N、C/P和N/P比的影响 Fig. 3 Effect of crabs on soil C/N, C/P and N/P ratios |
| 表 2 C/N、C/P和N/P比重复测量方差分析 Table 2 Repeated measure analysis of variance of C/N, C/P and N/P ratios |
ZP组土壤C/N比在7.89~9.11之间, 平均值为8.57±0.27, 变异系数为3.18%;C/P比在15.72~21.1之间, 平均值为18.41±0.68, 变异系数为3.71%;N/P比在1.99~2.27之间, 平均值为2.11±0.18, 变异系数为8.6%;ZP组土壤C/N比和C/P比相对稳定, 而N/P比变化较大.ZD组土壤C/N比在8.04~9.48之间, 平均值为8.52±0.35, 变异系数为4.14%;C/P比在15.02~18.54之间, 平均值为16.72±0.32, 变异系数为1.93%;N/P比在1.85~2.22之间, 平均值为2.01±0.19, 变异系数为9.39%;ZD组土壤C/N比和C/P比相对稳定, 而N/P比变化较大.在中潮滩中, 螃蟹作用下3种比值均大于对照组, 土壤C/N比和C/P比与不同季节呈显著关系(p<0.001), 而N/P比与不同季节呈较显著关系(p<0.05), 另外, 土壤C/P和N/P比分别与螃蟹作用关系极显著(p<0.001)和较为显著(p<0.05).
3.4 环境因子对土壤碳、氮、磷含量及其生态化学计量比的影响在不同潮滩, 土壤碳、氮、磷含量变化影响因子差异明显(表 3).GP组土壤碳含量与pH存在显著的正相关关系(p<0.05), 土壤磷含量与EC存在显著的负相关关系(p<0.05);ZP组土壤磷含量与土壤pH存在显著的正相关关系(p<0.05);ZD组土壤碳含量和氮含量与土壤容重存在显著的负相关关系(p<0.05).综合来看, 土壤碳含量受pH、EC、容重和含水量等多因子影响, 且土壤EC和容重与土壤碳、氮、磷含量变化的影响最大.
| 表 3 土壤碳、氮、磷含量与环境影响因子间的相关关系(n=12) Table 3 Correlation between TC, TN and TP concentration and environmental influencing factors |
从土壤理化性质的角度分析土壤C/N、C/P、N/P比的影响因素, 如各自比例、土壤pH和含水量等均存在一定的影响(表 4).GP组土壤磷含量对土壤C/N比有重要的影响, 存在显著的负相关关系(p<0.05);ZP组土壤N/P比与土壤pH存在显著的负相关关系(p<0.05), 土壤C/P比与土壤含水量存在显著的正相关关系(p<0.05);ZD组土壤C/P比与土壤含水量存在显著的正相关关系(p<0.01), 土壤C/N比与土壤含水量存在显著的正相关关系(p<0.05).综合来看, 土壤C/N和C/P比与土壤含水量存在显著正相关关系(p<0.05).
| 表 4 C/N、C/P和N/P比与土壤碳、氮、磷及环境影响因子间的相关关系(n=12) Table 4 Correlation between C/N, C/P and N/P ratios, and TC, TN and TP concentration, environmental influencing factors |
螃蟹影响下土壤碳、氮、磷储量的季节变化见图 4, GP组碳储量在2423.99~2791.71 t·km-2之间, 平均值为(2635.32±247.25) t·km-2, GD组碳储量在2168.43~2862.34 t·km-2之间, 平均值为(2539.79±209.34) t·km-2, GP组碳储量显著高于GD组碳储量(p<0.05);而ZP组碳储量在2089.53~2625.21 t·km-2之间, 平均值为(2352.90±173.14) t·km-2, ZD组碳储量在2510.72~2617.81 t·km-2之间, 平均值为(2577.06±128.85) t·km-2, ZP组碳储量显著低于ZD组碳储量(p<0.05), 说明在高潮滩和中潮滩有螃蟹活动下土壤碳储量相对较高, 变化较为明显.
 |
| 图 4 螃蟹对土壤碳、氮和磷储量的影响 Fig. 4 Effect of crabs on the soil carbon, nitrogen and phosphorus content |
GP组氮储量在263.76~331.50 t·km-2之间, 平均值为(300.54±25.27) t·km-2, GD组氮储量在261.81~329.47 t·km-2之间, 平均值为(291.64±23.72) t·km-2, GP组与GD组氮储量差异不显著(p>0.05);而ZP组氮储量在253.48~311.04 t·km-2之间, 平均值为(280.37±25.74) t·km-2, ZD组氮储量在275.52~323.07 t·km-2之间, 平均值为(298.80±14.73) t·km-2, ZP组氮储量显著低于ZD组氮储量(p<0.05), 说明在中潮滩有螃蟹活动下土壤氮储量相对较高, 变化较为明显.
GP组磷储量在126.68~160.86 t·km-2之间, 平均值为(139.22±11.81) t·km-2, GD组磷储量在131.69~164.09 t·km-2之间, 平均值为(143.66±15.57) t·km-2, GP组与GD组磷储量差异不显著(p>0.05);而ZP组磷储量在115.25~156.43 t·km-2之间, 平均值为(133.53±10.29) t·km-2, ZD组磷储量在134.33~174.44 t·km-2之间, 平均值为(150.19±10.95) t·km-2, ZP组磷储量显著低于ZD组磷储量(p<0.05).
土壤养分储量与C/N、C/P、N/P比关系如表 5所示, GD组磷储量与土壤N/P比呈显著负相关(p<0.05), ZP组磷储量与土壤C/P比呈显著负相关(p<0.05), ZD组氮储量与土壤C/N比呈显著负相关(p<0.05), 磷储量与土壤C/P比(p<0.05)和土壤N/P比(p<0.01)呈显著负相关关系, 综合来看, 土壤磷储量与C/N、C/P、N/P比关系密切.
| 表 5 土壤碳、氮和磷储量与C/N、C/P和N/P的相关关系(n=12) Table 5 Correlation between soil carbon, nitrogen and phosphorus reserves and C/N, C/P and N/P ratios |
土壤养分代谢过程及其环境响应是生态系统研究的重要方向(Liu et al., 2017), 河口湿地土壤营养元素C、N、P含量变化更具复杂性, 受到微生物活动、动物干扰、动植物残体腐解、潮汐运动和人类活动干扰等影响(刘文龙等, 2014).本研究中, 螃蟹干扰下土壤碳、氮、磷含量差异明显, 存在着明显的季节变化特征, 土壤碳、氮、磷含量最高出现在秋季, 主要是因为在秋季植物逐渐枯萎, 大量的凋落物输入到土壤中, 随着凋落物的分解过程, 增加了土壤中碳氮、磷含量; 与此同时, 在秋季温度相对夏季较低, 随着温度的降低碳、氮、磷元素的矿化速率也相对减弱, 从而促进碳、氮、磷元素在土壤中的积累(尹晓敏等, 2010; Liu et al., 2017).螃蟹可通过挖洞活动直接干扰土壤性质(酸碱度、通气性、颗粒组成等)(Simioni et al., 2014).本研究中, 在螃蟹的活动下, 土壤全碳和全氮含量高于对照组.主要与以下几方面的原因有关(Reinsel, 2004; Botto et al., 2006):①土壤动物通过摄食和消化作用将食物转化, 并以排泄物的形式将代谢产物排入到土壤中, 增加土壤有机质的输入量, 从而增加土壤中养分含量, 特别是碳、氮含量(Rickover et al., 2015; Li et al., 2016); ②土壤动物活动可以促进枯落物的碎化与分解, 提高枯落物分解过程向土壤中的有机质输入, 显著促进土壤中碳和养分的积累(Werry et al., 2005; Chen et al., 2016); ③土壤动物死亡后其残体也是其生境土壤碳与养分的来源, 随着土壤动物残体分解过程也会提高土壤中碳与养分的积聚; ④随着土壤动物活动的增加, 土壤中氧化环境增强, 使得土壤铁类物质的氧化物增多, 从而强化铁氧化物结合态碳的固定与稳定.与此同时, 本研究中螃蟹并未对土壤中磷的积聚起到促进作用, 主要是因为河口湿地磷的来源途径较为广泛(李洁等, 2015; 陈为峰等, 2008), 包括动植物残体输入、河水中营养物质磷输入、大气干湿沉降等.这种多来源途径使得磷的代谢过程较为复杂(Zhang et al., 2012), 也导致土壤磷的含量在螃蟹干扰下并未发生显著的增加, 这与Mortimer等(1999)的研究结果一致.
4.2 螃蟹影响下土壤碳、氮、磷生态化学计量学特征及其指示作用受土壤动物、植被、气候等多个土壤因子及人类干扰, 土壤碳、氮、磷含量也将发生变化, 呈时空变异性(Cleveland et al., 2007; 刘文龙等, 2014).本研究发现, 螃蟹干扰下土壤C/N、C/P、N/P比在不同季节有不同, 夏季出现低峰值(图 3).可能是因为在夏季温度较高, 微生物活性也较高, 土壤碳的释放量也相对较大, 从而降低在元素比值中的碳含量和碳与养分比值, 同时也说明土壤碳与养分比值受土壤碳固持与输入的控制(肖烨等, 2014); 对于氮磷比值的降低, 主要是因为相对于氮养分而言, 磷的矿化与淋溶损失的效率较慢, 从而使得在夏季高温季节中, 氮磷比值较低.与此同时, 从本研究中养分相关性的分析结果发现, 氮对环境变化的响应也快于磷, 这与前人研究相似(Cleveland et al., 2007; 王维奇等, 2010), 也从另一个侧面解释了在夏季高温季节氮磷比值相对较低的原因.此外, 在闽江河口湿地, 互花米草生长过程中存在着明显的氮限制, 在其生长旺盛的夏季, 其对氮的吸收量将会增加, 从而降低土壤中的氮含量, 从而形成较低的氮磷比.此外, 本研究中, 螃蟹干扰下的土壤C/N、C/P、N/P比值大于无螃蟹作用下的相应比值, 土壤C/N、C/P、N/P比均表现为GP>ZP和GD>ZD, 这种空间的变异性主要受控于碳、氮、磷养分积聚在空间上变化的规律.
此外, 生态化学计量学特征也具有重要的指示作用, 其中, 碳与养分比值对碳储量具有重要的指示意义.本研究发现, 从总体上来看, 生态化学计量学特征比值与碳储量存在显著的正相关关系, 特别是碳与养分比值, 这与王维奇等(2010)的研究结论相似.与此同时, 本研究区螃蟹作用下的土壤N/P比(2.14)低于全国湿地N/P比(13.60)(张仲胜等, 2016).一般而言, 较低的N/P比指示着较高的初级生产力; 其次, 植物的地上部呼吸作用受氮素营养的影响, 而螃蟹活动加强了氮素的转移, 有助于植株地上部的呼吸, 结合本研究区所处亚热带雨热同期的气候背景下较高初级生产力的现状, 间接说明氮养分可能是植物生长过程中的潜在限制性养分(王维奇等, 2010).
5 结论(Conclusions)1) 湿地土壤碳、氮、磷含量具有较为明显的季节变化, 且在高、中潮滩有所不同, 螃蟹影响提高了湿地土壤碳、氮含量, 但并未促进土壤磷含量的累积.
2) 螃蟹影响下土壤碳和氮、氮与磷元素之间呈现良好的线性拟合关系, 而碳与磷元素之间线性拟合的程度相对较低; 螃蟹影响下生态化学计量学特征对土壤碳储量和植物限制性养分指示作用不强.
3) 湿地土壤碳、氮、磷生态化学计量学特征具有较为明显的季节变化, 且在高、中潮滩有所不同, 螃蟹活动一定程度上增加了土壤C/N、C/P、N/P比值, 但影响并不显著.
Allen A P, Gillooly J F. 2009. Towards an integration of ecological stoichiometry and the metabolic theory ecology to better understand nutrient cycling[J]. Ecology Letters, 12(5): 369–384.
DOI:10.1111/ele.2009.12.issue-5
|
Botto F, Iribarne O, Gutierrez J, et al. 2006. Ecological importance of passive deposition of organic matter into burrows of the SW Atlantic crab Chasmagnathus granulatus[J]. Marine Ecology Progress Series, 312(2): 1–10.
|
Chen G C, Lu C, Li R, et al. 2016. Effects of foraging leaf litter of Aegiceras corniculatum (Ericales, Myrsinaceae) by Parasesarma plicatum (Brachyura, Sesarmidae) crabs on properties of mangrove sediment:a laboratory experiment[J]. Hydrobiologia, 763(1): 125–133.
DOI:10.1007/s10750-015-2367-1
|
陈为峰, 史衍玺, 田素锋, 等. 2008. 黄河口新生湿地土壤氮磷分布特征研究[J]. 水土保持学报, 2008, 22(1): 69–73.
|
Cleveland C C, Liptzin D. 2007. C:N:P stoichiometry in soil:is there a " Redfield ratio " for the microbial biomass?[J]. Biogeochemistry, 85(3): 235–252.
DOI:10.1007/s10533-007-9132-0
|
Elser J J, Stemer R M, Gorokhova E, et al. 2000. Biological stoichiometry from genes to ecosystems[J]. Ecology Letters, 3(6): 540–550.
DOI:10.1046/j.1461-0248.2000.00185.x
|
Elser J J, Bracken M E S, Cleland E E. 2007. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems[J]. Ecology Letters, 10(12): 1135–1142.
DOI:10.1111/ele.2007.10.issue-12
|
Han W, Fang J Y, Guo D, et al. 2005. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J]. New Phytologist, 168(2): 377–385.
DOI:10.1111/j.1469-8137.2005.01530.x
|
Hu Z, Wu S, Ji C, et al. 2016. A comparison of methane emissions following rice paddies conversion to crab-fish farming wetlands in southeast China[J]. Environmental Science and Pollution Research, 23(2): 1505–1515.
DOI:10.1007/s11356-015-5383-9
|
胡知渊, 鲍毅新, 程宏毅, 等. 2009. 中国自然湿地底栖动物生态学研究进展[J]. 生态学杂志, 2009, 28(5): 959–968.
|
李洁, 张文强, 金鑫, 等. 2015. 环渤海滨海湿地土壤磷形态特征研究[J]. 环境科学学报, 2015, 35(4): 1143–1151.
|
Li S, Zhang S, Pu Y, et al. 2016. Dynamics of soil labile organic carbon fractions and C-cycle enzyme activities under straw mulch in Chengdu Plain[J]. Soil and Tillage Research, 155: 289–297.
DOI:10.1016/j.still.2015.07.019
|
刘剑秋, 曾从盛, 陈宁. 2006. 闽江河口湿地研究[M]. 北京: 科学出版社: 330–334.
|
刘敏, 侯立军, 许世远, 等. 2005. 长江口潮滩生态系统氮微循环过程中大型底栖动物效应实验模拟[J]. 生态学报, 2005, 25(5): 1132–1137.
|
刘文龙, 谢文霞, 赵全升, 等. 2014. 胶州湾芦苇潮滩土壤碳、氮和磷分布及生态化学计量学特征[J]. 湿地科学, 2014, 12(3): 362–368.
|
Liu Y, Jiang M, Lu X, et al. 2017. Carbon, nitrogen and phosphorus contents of wetland soils in relation to environment factors in northeast China[J]. Wetlands, 37(1): 153–161.
DOI:10.1007/s13157-016-0856-2
|
鲁如坤. 2000. 土壤农业化学分析方法[M]. 北京: 中国农业科学出版社.
|
Mortimer R J G, Davey J T, Krom M D, et al. 1999. The effect of macrofauna on porewater profiles and nutrient fluxes in the intertidal zone of the humber estuary estuarine coastal and shelf[J]. Science, 48(6): 683–699.
|
Reinsel K A. 2004. Impact of fiddler crab foraging and tidal inundation on an intertidal sand flat:season-dependent effects in one tidal cycle[J]. Journal of Experimental Marine Biology and Ecology, 313(1): 1–17.
DOI:10.1016/j.jembe.2004.06.003
|
Rickover L, Khabarovsk T, Howe A, et al. 2015. Microbial activity in forest soil reflects the changes in ecosystem properties between summer and winter[J]. Environmental Microbiology, 18(1): 288–301.
|
Simioni F, Campos V A, Dorado-Rodrigues T F, et al. 2014. Crab burrows and termite thermal chimneys as refuges for anurans in a Neotropical wetland[J]. Salamandra, 50(3): 133–138.
|
Stahl M O, Tarek M H, Yeo D C J, et al. 2014. Crab burrows as conduits for groundwater-surface water exchange in Bangladesh[J]. Geophysical Research Letters, 41(23): 8342–8347.
DOI:10.1002/2014GL061626
|
Tessier J T, Raynal D J. 2003. Use of nitrogen to phosphorus ratio in plant tissue as indicator of nutrient limitation and nitrogen saturation[J]. Journal of Applied Ecology, 40(3): 523–534.
DOI:10.1046/j.1365-2664.2003.00820.x
|
Vermeiren P, Sheaves M. 2015. Modeling intertidal crab distribution patterns using photographic mapping among tropical Australian estuaries[J]. Estuaries and Coasts, 38(5): 1545–1556.
DOI:10.1007/s12237-014-9897-z
|
王绍强, 于贵瑞. 2008. 生态系统碳氮磷元素的生态化学计量学特征[J]. 生态学报, 2008, 28(8): 3937–3947.
|
王维奇, 曾从盛, 钟春棋, 等. 2010. 人类干扰对闽江河口湿地碳、氮、磷生态化学计量学特征的影响[J]. 环境科学, 2010, 31(10): 2411–2416.
|
王霞, 李辉信, 朱玲, 等. 2008. 蚯蚓活动对土壤氮素矿化的影响[J]. 土壤学报, 2008, 45(4): 641–648.
|
Werry J, Lee S Y. 2005. Grapsid crabs mediate link between mangrove litter produetion and estuarine planktonic food chains[J]. Marine Ecology Progress Series, 293: 165–176.
DOI:10.3354/meps293165
|
肖烨, 薛振山, 张仲胜, 等. 2014. 吉林东部山地沼泽湿地土壤碳、氮、磷含量及其生态化学计量学特征[J]. 地理科学, 2014, 34(8): 994–1001.
|
Yang Y H, Fang J Y, Guo D L, et al. 2010. Vertical patterns of soil carbon, nitrogen and carbon:nitrogen stoichiometry in Tibetan grasslands[J]. Biogeosciences Discussions, 7(1): 1–24.
DOI:10.5194/bgd-7-1-2010
|
尹晓敏, 吕宪国, 等. 2010. 湿地土壤Fe与N耦合过程研究进展[J]. 环境科学, 2010, 31(9): 2254–2259.
|
张仲胜, 吕宪国, 薛振山, 等. 2016. 中国湿地土壤碳氮磷生态化学计量学特征研究[J]. 土壤学报, 2016, 53(5): 1160–1169.
|
Zhang Z S, Lu X G, Song X L, et al. 2012. Soil C, N and P stoichiometry of Deyeuxia angustifolia and Carex lasiocarpa wetlands in Sanjiang Plain, Northeast China[J]. Journal of Soils and Sediments, 12(9): 1309–1315.
DOI:10.1007/s11368-012-0551-8
|
曾从盛, 钟春棋, 仝川, 等. 2008. 土地利用变化对闽江河口湿地表层土壤有机碳含量及其活性的影响[J]. 水土保持学报, 2008, 22(5): 125–129.
|
Zhou J, Wu Y, Kang Q, et al. 2007. Spatial variations of carbon, nitrogen, phosphorous and sulfur in the salt marsh sediments of the Yangtze Estuary in China[J]. Estuarine, Coastal and Shelf Science, 71: 47–59.
DOI:10.1016/j.ecss.2006.08.012
|