 2018, Vol. 38
2018, Vol. 38



2. 甘肃农业大学林学院, 兰州 734000
2. College of Forestry, Gansu Agricultural University, Lanzhou 734000
大气中挥发性有机物(Volatile Organic Compounds, 简写为VOCs)中的很多成分具有高度的化学活性, 极易与大气中的NO、NO2、氧化剂和·OH等发生反应(唐孝炎等, 2010;Fehsenfeld et al., 1992; Laothawornkitkul et al., 2009), 在对流层O3光化学过程中具有重要的作用(Possell et al., 2011; Monson et al., 2001).另外, VOCs为形成O3的重要前体物之一, 对气候变暖有间接影响(Collins et al., 2002; Millet et al., 2016).准确确定VOCs排放特征及影响因素, 对科学认识对流层O3光化学过程及大气环境变化和气候变暖都有重要的理论意义.大气VOCs中非甲烷碳氢化合物(Non-methane Hydrocarbon, 简写为NMHC)总量的90%以上来自于陆地生物圈排放, 异戊二烯(Isoprene, 分子式为C5H8)是对流层中最主要的非甲烷类碳氢化合物, 其主要来源于植物排放, 排放量约占植物非甲烷碳氢化合物排放总量的44%(Guenther et al., 1995), 也是农村和边远地区大气中浓度最高的非甲烷类碳氢化合物(Fehsenfeld et al., 1992), 对城市大气中的光化学过程和大气环境影响也较大(Possell et al., 2011; Monson et al., 2001).植物排放异戊二烯过程受植物生理活动、土壤水分和养分、温湿度、降水与蒸发、太阳辐射等的影响, 但光合有效辐射和温度是最主要的影响因子(Monson et al., 2001; Potter et al., 2001).植物排放异戊二烯的速度随叶片温度和光合光量子密度的增加而增加(Monson et al., 2001).随着全球气候变化, 叶温和光合有效辐射光量子密度将改变, 进而将使植物异戊二烯排放速率改变, 这对O3光化学过程及大气环境将产生一定的影响(Laothawornkitkul et al., 2009; Turner et al., 1991; Zhang et al., 2008).因此, 准确确定气候变化对植物异戊二烯排放速率的影响, 对科学认识气候变化对O3光化学过程及大气环境影响等具有重要的理论和现实意义.
国际上已广泛展开了对植物源挥发性有机物(BVOCs)排放的研究, 包括测定不同植物异戊二烯排放速率(Harley et al., 2004), 计算植物异戊二烯排放清单(Potter et al., 2001), 分析过去植物异戊二烯排放(Unger, 2013)和全球植被BVOCs排放量(Levis et al., 2003; Lathière et al., 2005), 以及分析光照(Laffineur et al., 2013)、温度和O3浓度(Hartikainen et al., 2012)及水分(Copolovici et al., 2010)变化对植物BVOCs排放的影响, 模拟不同温度下植物异戊二烯排放速率等(Monson et al., 2012).随着气候变化与大气环境关系研究的深入, 有关气候变化对植物异戊二烯排放影响的研究也倍受关注, 包括模拟气候变化对植物异戊二烯排放的影响(Constable et al., 1999), 观测气候变化对植物BVOCs排放的影响(Rinnan et al., 2011; Kramshøj et al., 2016), 分析气候变化对植物组成和异戊二烯排放的影响(Kellomäki et al., 2001), 干旱对植物异戊二烯排放的影响(Pegoraro et al., 2006; Peñuelas et al., 2009), CO2浓度变化对植被BVOCs排放的影响(Possell et al., 2011)及温度和CO2浓度变化(Potosnak et al., 2014)或O3和CO2浓度变化(Calfapietra et al., 2007)对植物异戊二烯排放的影响等, 以及综合分析土地利用、CO2浓度、植被组成和气候变化对森林异戊二烯排放的影响(Arneth et al., 2008)或全球变化对植物BVOCs排放的影响(Monson et al., 2007)等.这些研究极大地提高了人们对气候变化对植物异戊二烯排放影响的认识, 但研究主要集中在欧洲和北美或高纬度地区, 并且主要是针对气候变化对植物异戊二烯排放短期影响观测分析或植被组成变化对异戊二烯排放影响分析等, 对未来气候变化对其它地区植物异戊二烯排放影响的研究还不足.我国也有许多对植物BVOCs排放的研究报道, 包括测定植物异戊二烯排放速率(白建辉等, 2012;2015;陈颖等, 2009;梁宝生等, 2003), 分析植物BVOCs排放速率(王志辉等, 2003)、植物排放BVOCs组成(梁珍海等, 2011)和释放规律(宁平等, 2013)等, 以及测定珠江三角洲(司徒淑娉等, 2009)、西双版纳(王志辉等, 2003)和太湖地区(王效科等, 2002)典型植物异戊二烯排放特征等.另外, 也进行了森林异戊二烯排放(张莉等, 2003)和植被BVOCs排放清单分析(闫雁等, 2005)等.这些研究提高了人们对植物异戊二烯排放特征的认识, 但有关气候变化对植物异戊二烯排放影响的研究还未见报道.
我国亚热带植被分布广泛.毛竹(Phyllostachys pubescens)是亚热带植被中典型的多年生常绿树种, 其异戊二烯排放速率高(周秀骥等, 2004;白建辉等, 2015).目前, 已有学者对毛竹的异戊二烯排放进行了观测(周秀骥等, 2004;白建辉等, 2015), 但有关气候变化对这种植物异戊二烯排放的影响研究还未见报道.为此, 本文采用全球气候模式NorESM1-M产生的RCP2.6、RCP4.5、RCP6.0和RCP8.5气候变化情景数据和植物异戊二烯排放计算模型, 模拟分析未来气候变化对毛竹叶片异戊二烯排放速率的影响, 以期为科学认识气候变化对植物BVOCs排放的影响提供依据, 并为相关研究提供一定的参考.
2 材料与方法(Materials and methods) 2.1 毛竹异戊二烯排放计算参数基于文献(周秀骥等, 2004), 确定毛竹在标准状态下(T=303 K, 光合有效辐射量子密度PAR=1000 μmol·m-2·s-1)的异戊二烯排放速率为116.04 μg·g-1·h-1.毛竹分布广泛, 为集中分析气候变化对毛竹异戊二烯排放速率的影响, 本文以毛竹分布集中区域——武夷山自然保护区(27.55°~27.90°N, 117.45°~117.85°E)为例, 该区域处在中亚热带温暖湿润季风气候区, 而对气候变化下不同分布区毛竹异戊二烯排放速率差异比较将另文介绍.
2.2 毛竹异戊二烯排放速率的计算参考文献(Guenther et al., 1993), 遵循植物异戊二烯排放速率计算的一般模型, 采用公式(1)计算毛竹的异戊二烯排放通量.
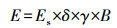
|
(1) |
式中, E为异戊二烯排放通量(μg·g-1·h-1);Es为标准条件下(T=303 K, 光合有效辐射量子密度PAR=1000 μmol·m-2·s-1)毛竹的异戊二烯排放量(μg·g-1·h-1);δ为考虑长期排放活性的控制因子, 由于毛竹为常绿植物, 所以δ取值为1;B为叶片生物量密度, 由于本文主要分析植物的异戊二烯排放速率, 所以单位生物量质量取值为1;γ为活性排放因子, 为叶温和光合有效辐射的函数, 具体利用公式(2)~(4)进行计算.
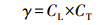
|
(2) |
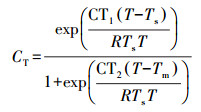
|
(3) |
式中, CT1取值为95000 J·mol-1, T为植物叶片的温度(K), Ts为标准条件下(PAR=1000 μmol·m-2·s-1)下叶片温度(303 K), Tm取值为314 K, CT2取值为23000 J·mol-1, R为气体常数(8.314 J·K-1·mol-1).
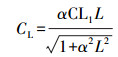
|
(4) |
式中, α为常数(0.0027), CL1取值为1.066, L为每小时光合有效辐射量子密度(μmol·m-2·s-1).
以上公式计算获得的毛竹异戊二烯排放速率为逐时值, 且叶温和光合有效辐射量子密度也都是逐时值.由于缺乏叶温观测资料, 而用能量守衡方法计算叶温又非常复杂, 本文参照文献(张莉等, 2003;周秀骥等, 2004)中的方法, 直接用气温代替叶温计算叶片温度逐时值.每小时光合有效辐射量子密度按每小时辐射强度值经过转换计算.计算温度和光合有效辐射量子密度逐时值的具体方法见2.3.2节和2.3.3节.
2.3 气候变化影响下毛竹异戊二烯排放速率计算 2.3.1 气候变化情景数据采用全球气候模式NorESM1-M产生的RCP2.6、RCP4.5、RCP6.0和RCP8.5情景数据(RCP是Representative Concentration Pathways简写, RCP2.6、RCP4.5、RCP6.0和RCP8.5分别代表大气中辐射强迫分别为2.6、4.5、6.0和8.5 W·m-2).在RCP2.6情景中, 未来人类将采取大力减排措施, 使辐射强迫在21世纪达到顶峰并下降, 到2100年大气中CO2浓度将达到421 ×10-6, 辐射强迫高达2.6 W·m-2, 全球升温幅度将达0.3~1.7 ℃;RCP8.5情景是最高的温室气体排放情景, 假定人口增长最快, 技术革新水平较低, 能源改善缓慢, 导致持续过高的能源需求及大量温室气体排放, 到2100年大气中CO2浓度将达936 ×10-6, 辐射强迫将持续上升, 到2100年将高达8.5 W·m-2, 全球升温幅度将达到2.6~4.8 ℃;RCP4.5和RCP6.0情景介于RCP2.6和RCP8.5情景之间, 在RCP4.5情景下2100年辐射强迫高达4.5 W·m-2, 升温幅度将达1.1~2.6 ℃, 在RCP6.0情景下2100年辐射强迫达6.0 W·m-2, 升温幅度达1.4~3.1 ℃(Taylor et al., 2012; Meinshausen et al., 2011; Warszawaski et al., 2013; Hempel et al., 2013).这些情景数据是由中国科学院地理与资源研究所从ISI-MIP(the inter-sectional impact model inter-comparison project, 多领域间影响模型比较计划)获取的经过空间降尺度、偏差校正的RCP情景下的全球气候模式模拟数据, 由中国农业科学院进行订正, 在本课题组承担的国家“十二五”科技支撑课题中已使用(吴建国, 2017).
2.3.2 温度日变化计算气候变化情景中温度是逐日值, 在计算毛竹的异戊二烯排放中需要按小时值进行计算, 所以需要把温度日值转化为逐时值.参考文献(Ephrath et al., 1996), 白天温度逐时值Ta按公式(5)计算.
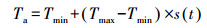
|
(5) |
式中, Tmin为最低温度(K), Tmax为最高温度(K), s(t)为计算温度变化函数, 按照公式(6)计算.
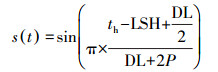
|
(6) |
式中, π为常数;th为一天太阳时(h), 按照公式(7)计算;LSH为最大太阳高度时刻, 取值为12 h;DL为白昼长度(h), 按照公式(10)计算;P为最高气温与最大太阳高度出现的时差, 取值2.
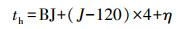
|
(7) |
式中, BJ为北京时(h);J为地理经度(°);η为时差(h), 参考文献(盛裴轩等, 2003), 按照公式(8)计算.
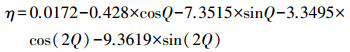
|
(8) |
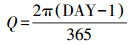
|
(9) |
式中, DAY为1年中日序, 每年1月1日DAY=1, 1月2日DAY=2, 依次类推, 12月31日DAY=365.
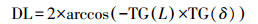
|
(10) |
式中, L为地理纬度(°);δ为太阳赤纬(rad), 参考文献(盛裴轩等, 2003), 按照公式(11)计算.
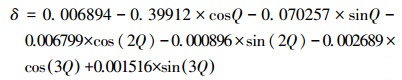
|
(11) |
白天时刻按照公式(12)判断, 夜间时刻按照公式(13)判断.夜间每小时温度值按照公式(14)计算.
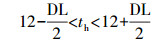
|
(12) |

|
(13) |
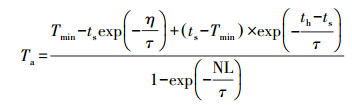
|
(14) |
式中, Tmin为日最低气温(K);th为一天太阳时(h), 按公式(7)计算;ts为日落时间, 取值为LSH+DL/2;NL为夜间长度, 取值NL=24-DL;τ为时间系数, 取值4 h.
2.3.3 光合有效辐射量子密度日变化计算计算毛竹异戊二烯排放速率时需要先计算光合有效辐射量子密度的逐时值.气候变化情景中辐射值为日总辐射值, 需要把辐射日值转换为光合有效辐射量子密度逐时值.参考文献(Ephrath et al., 1996), 按照公式(15)计算每小时光合有效辐射强度, 再参考文献中方法(周允华等, 1996), 计算每小时光合有效辐射量子密度值.
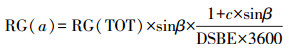
|
(15) |
式中, RG(α)为每个时刻的太阳辐射中光合有效辐射强度(W·m-2), RG(TOT)为总太阳辐射中光合有效辐射强度(W·m-2);c为气象常数, 取值0.4;sinβ为太阳高度正弦函数, 按照式(16)计算;DSBE为sinβ×sin(1+csinβ)从太阳升起到太阳落山时段的每天积分, 按照式(19)计算.
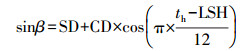
|
(16) |
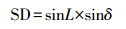
|
(17) |
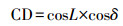
|
(18) |
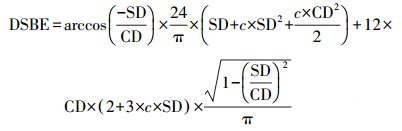
|
(19) |
式中, SD为太阳高度正弦函数季节补偿因子;CD为太阳高度正弦函数正幅因子;th为一天太阳时(h), 按照式(12)计算;LSH为最大太阳高度出现时刻, 取值12 h;c为常数, 一般取0.4.
2.3.4 毛竹异戊二烯排放速率计算把基准和未来气候变化情景下温度和光合有效辐射量子密度逐时值, 代入式(1)~(4)计算毛竹异戊二烯逐时排放速率.在此基础上, 进行24 h累加, 计算毛竹异戊二烯逐日排放速率;基于毛竹异戊二烯逐日排放速率, 再经过周年累加, 计算毛竹异戊二烯逐年排放速率.通过比较基准和未来气候变化情景下毛竹异戊二烯日排放和年排放速率差异, 分析气候变化对毛竹异戊二烯日排放速率和年排放速率的影响.以上计算分析都在Visual Fortran6.6平台程序中进行.
3 结果(Results) 3.1 气候变化特征图 1~3是根据气候变化情景数据计算的武夷山自然保护区范围的气温、降水量和辐射变化特征.图 1显示, 相比1951—2000年基准情景, 2001—2050年武夷山自然保护区年均气温呈显著上升趋势(p<0.05).相比1951—2000年, 在2001—2050年时段, 武夷山自然保护区年均气温在RCP2.6、RCP4.5、RCP6.0和RCP8.5情景下分别上升0.90、0.95、0.97和1.14 ℃(图 1).相比1951—2000年, 在2001—2025年时段, 武夷山自然保护区年均气温在RCP2.6、RCP4.5、RCP6.0和RCP8.5情景下分别上升0.60、0.46、0.63和0.57 ℃;到2026—2050年, 武夷山自然保护区年均气温在RCP2.6、RCP4.5、RCP6.0和RCP85情景下分别上升1.19、1.44、1.31和1.70 ℃.
 |
| 图 1 相比基准情景(1951—2000年) 2001—2050年平均气温变化趋势 Fig. 1 Changes in air temperature in 2001—2050 compared with baseline periods(1951—2000) |
图 2显示, 相比1951—2000年基准情景, 2001—2050年时段武夷山自然保护区年降水量呈较大波动, 在RCP2.6、RCP4.5、RCP6.0和RCP8.5情景下变化不同.相比1951—2000年, 2001—2050年时段, 武夷山自然保护区年降水量在RCP2.6、RCP4.5、RCP6.0和RCP8.5情景下分别增加58.04 mm、增加64.47 mm、减少69.09 mm和增加35.03 mm(图 2).相比1951—2000年时段, 2001—2025年时段, 武夷山自然保护区年降水量在RCP2.6、RCP4.5、RCP6.0和RCP8.5情景下分别减少50.64 mm、增加67.14 mm、减少56.04 mm和减少57.87 mm;2026—2050年, 武夷山自然保护区年降水量在RCP2.6、RCP4.5、RCP6.0和RCP8.5情景下分别增加166.73 mm、增加61.81 mm、减少82.13 mm和增加127.94 mm.
 |
| 图 2 相比基准情景(1951—2000年) 2001—2050年年降水量的变化趋势 Fig. 2 Changes in annual preciptation in 2001—2050 compared with baseline periods(1951—2000) |
图 3显示, 相比1951—2000年基准情景, 2001—2050年时段武夷山自然保护区辐射强度呈较大波动.相比1951—2000年基准情景, 2001—2050年时段武夷山自然保护区辐射强度在RCP2.6、RCP4.5、RCP6.0和RCP8.5情景下分别减少0.104、0.232、0.207和0.118 MJ·m-2.相比1951—2000年基准情景, 2001—2025年时段武夷山自然保护区辐射强度在RCP2.6、RCP4.5、RCP6.0和RCP8.5情景下分别减少0.366、0.558、0.271和0.241 MJ·m-2;2026—2050年时段, 武夷山自然保护区辐射强度在RCP2.6、RCP4.5、RCP6.0和RCP8.5情景下分别增加0.157 MJ·m-2、增加0.094 MJ·m-2、减少0.143 MJ·m-2和增加0.006 MJ·m-2.
 |
| 图 3 相比基准情景(1951—2000年) 2001—2050年太阳总辐射变化趋势 Fig. 3 Changes in total solar radiation in 2001—2050 compared with baseline periods(1951—2000) |
图 4是1951—2000年基准情景和2001—2050年RCP2.6、RCP4.5、RCP6.0和RCP8.5情景下50年平均的毛竹异戊二烯日排放速率值.结果显示, 在基准情景、RCP2.6、RCP4.5、RCP6.0和RCP8.5情景下, 毛竹异戊二烯日排放速率平均值分别为371、406、407、406和419 μg·g-1·d-1;毛竹异戊二烯日排放速率平均值在未来气候变化情景下比基准情景下高约30 μg·g-1·d-1, 在RCP8.5情景下最高.但在基准情景、RCP2.6、RCP4.5、RCP6.0和RCP8.5情景下, 毛竹异戊二烯在不同日期的日排放速率变化很大.在1~210 d毛竹异戊二烯日排放速率呈现上升趋势, 在210~365 d呈现下降趋势.按照对应月份, 毛竹异戊二烯日排放速率在5—9月较高, 在1—4月和9—12月较低.
 |
| 图 4 基准和不同气候变化情景下毛竹异戊二烯日排放速率 Fig. 4 Daily isoprene emission rate from leaves of Phyllostachys pubescens under baseline and climate change scenarios |
图 5显示, 毛竹异戊二烯日排放速率在RCP2.6、RCP4.5和RCP6.0情景下与基准情景下平均差异分别为35、36和35 μg·g-1·d-1, 在RCP8.5情景下与基准情景下平均差异为47 μg·g-1·d-1.总体上, 毛竹异戊二烯日排放速率在RCP2.6、RCP4.5和RCP6.0情景下与基准情景下差异在35 μg·g-1·d-1以上, 在RCP8.5情景下差异较大.但在RCP2.6、RCP4.5、RCP6.0和RCP8.5情景与基准情景下毛竹异戊二烯不同日期的差值变化很大.1~90 d, 未来气候变化情景与基准情景下毛竹异戊二烯日排放速率差值呈稳定趋势, 不同气候变化情景下与基准情景下毛竹异戊二烯日排放速率差异较小;91~300 d, 不同气候变化情景下与基准情景下毛竹异戊二烯日排放速率差值呈较大波动;301~365 d, 不同情景与基准情景下毛竹异戊二烯日排放速率差异不大.
 |
| 图 5 不同气候变化情景下与基准情景下毛竹异戊二烯日排放速率差异 Fig. 5 The difference of daily isoprene emission rate from leaves of Phyllostachys pubescens under baseline and climate change scenarios |
图 6是2001—2050年时段RCP2.6、RCP4.5、RCP6.0和RCP8.5情景下相比1951—2000年基准情景下50年平均的毛竹异戊二烯日排放速率相对变化百分率.图 6显示, 与基准情景下相比, 在RCP2.6、RCP4.5、RCP6.0和RCP8.5情景下, 在1~190 d, 毛竹异戊二烯日排放速率相对变化百分率较高(相比基准情景平均增加15%以上);在190~270 d, 毛竹异戊二烯日排放速率相对变化百分率较小;在271~365 d, 毛竹异戊二烯日排放速率相对变化百分率在20%.与基准情景比较, RCP2.6、RCP4.5、RCP6.0和RCP8.5情景下, 毛竹异戊二烯日排放速率变化幅度平均分别为12.53%、13.15%、13.05%和16.60%, 在RCP8.5情景下变化最大.
 |
| 图 6 不同气候变化情景下与基准情景下毛竹异戊二烯日排放速率变化率 Fig. 6 The change in daily isoprene emission rate from leaves of Phyllostachys pubescens under climate change scenarios compared to the baseline scenario |
图 7显示, 与基准情景下相比, 在RCP2.6、RCP4.5、RCP6.0和RCP8.5情景下, 2001—2050年毛竹异戊二烯年排放速率呈增加的趋势(p<0.05).在基准情景下毛竹异戊二烯年排放速率平均为135378 μg·g-1·a-1, 在RCP2.6、RCP4.5、RCP6.0和RCP8.5情景下毛竹异戊二烯平均年排放速率分别为147965、148549、147977和152615 μg·g-1·a-1.总体上, 毛竹异戊二烯年排放速率在未来气候变化情景下比基准情景下高10000 μg·g-1·d-1以上, 在RCP8.5情景下最大.
 |
| 图 7 不同气候变化情景下与基准情景下毛竹异戊二烯年排放速率变化 Fig. 7 Changes in annual emission rate of isoprene from leaves of Phyllostachys pubescens under different climate change scenarios compared to the baseline scenario |
图 7显示, 相比基准情景, 在RCP2.6、RCP4.5、RCP6.0和RCP8.5情景下, 2001—2050年毛竹异戊二烯年排放年速率变化程度呈增加趋势(p<0.05).相比基准情景, 在2001—2050年, 毛竹异戊二烯年排放速率在RCP2.6、RCP4.5、RCP6.0和RCP8.5情境下的平均增加幅度在9%~13%, 在RCP2.6情景下平均增加9.30%, 在RCP4.5情景下平均增加9.73%, 在RCP6.0情景下平均增加9.32%, 在RCP8.5情景下平均增加12.73%.总体上, 在RCP8.5情景下毛竹异戊二烯年排放速率变化幅度最大.
4 讨论(Discussion)本研究说明, 未来气候变化下武夷山自然保护区温度呈上升趋势, 这与全球趋势比较一致(Stocker et al., 2013).另外, 本研究也表明, 未来气候变化下武夷山自然保护区年降水量波动明显, 呈增加或下降趋势.同时, 气候变化下武夷山自然保护区辐射强度波动也比较大, 也呈增加或下降趋势, 这些趋势与全球气候变化预测中一致(Stocker et al., 2013).当然, 这里主要反映了武夷山自然保护区范围内的变化趋势, 不同区域将有不同的变化特征.
一些研究表明, 未来气候变化下将使植物异戊二烯排放速率增加.如Lindwall等(2016)发现, 夏天变暖4 ℃使极地冻原BVOCs排放增加280%, 积雪增加对BVOCs排放影响不大;Kramshøj等(2016)研究发现, 气候变暖使极地植物BVOCs排放增加, 变暖使生态系统BVOCs排放速率增加260%, 植物BVOCs排放速率增加90%, 减少太阳辐射强度使系统VOCs排放速率下降69%, 植物BVOCs排放速率下降61%~65%;Lindwall等(2016)研究发现, 气候变暖使极地沼泽BVOCs排放增加, 温度升高1.3~1.6 ℃下植物异戊二烯排放速率增加240%~380%;Faubert等(2012)研究发现, 气候变化使亚北极苔原BVOCs排放加倍;Rinnan等(2011)研究发现, 气候变化引起灌木BVOCs排放增加;Tiiva等(2008)研究发现, 气候变化使亚极地石楠中BVOCs排放增加;Turner等(1991)研究发现, 气候变化引起异戊二烯排放速率增加;Vedel-Petersen等(2015)研究发现, 气候变化导致植物BVOCs排放增加.还有研究发现, 温度变化使BVOCs排放增加(Filella et al., 2007), 区域气候变化使BVOCs排放增加(Zhang et al., 2008).Constable等(1999)研究发现, 气候变化并伴随CO2浓度加倍导致植物生物量增加, 使异戊二烯排放增加82%, 植被改变将使BVOCs排放减少, 使异戊二烯排放减少11%.本研究表明, 未来气候变化影响下毛竹异戊二烯日和年排放速率将明显改变, 并且在RCP8.5情景下改变程度最大, 同时存在年内时间差异.说明未来气候变化将使毛竹异戊二烯排放速率增加, 但一年中不同时间存在差异.这与一年中气候变化趋势及气候要素不同日期变化和植物生理活动时间差异都有关.气候变化下在冬、春季升温幅度相对较高, 夏季低, 但植物生理活动在冬季比较弱而在春、夏季节活跃, 导致气候变化影响下在一年中不同时段异戊二烯排放速率变化差异.白建辉等(2015)分析发现, 分布在浙江的竹林BVOCs排放主要集中于夏季(7—9月), 并且异戊二烯和单萜烯对各因子变化响应是非线性的, 当各个因子增加率大于40%时, 异戊二烯对光合有效辐射变化最敏感.同时, 气候变化下毛竹异戊二烯排放速率时间差异也与不同时间的气候变化情景下各个气候要素变化幅度有关.在未来气候变化下, 春、冬季温度变化幅度相对较大, 但绝对值仍旧没有夏季高, 辐射强度变化也存在时间差异, 导致在一年中不同时间毛竹异戊二烯排放速率变化不同.另外, 本研究也表明, 毛竹异戊二烯排放速率增加在RCP8.5情景下最高, 但在RCP6.0情景下没有比RCP4.5情景下高, 这与不同时间段及不同区域气候要素变化程度有关.在RCP2.6情景中, 未来人类将采取大力减排措施, 使辐射强迫在21世纪达到顶峰并下降, 到2100年大气中CO2浓度将达到421×10-6, 辐射强迫高达2.6 W·m-2, 全球升温幅度将达0.3~1.7 ℃;RCP8.5情景是最高的温室气体排放情景, 假定人口增长最快, 技术革新水平较低, 能源改善缓慢, 导致持续过高的能源需求及大量温室气体排放, 到2100年大气中CO2浓度将达936×10-6, 辐射强迫将持续上升, 到2100年将高达8.5 W·m-2, 全球升温幅度将达2.6~4.8 ℃;RCP4.5和RCP6.0情景介于RCP2.6和RCP8.5之间, 在RCP4.5情景下2100年辐射强迫高达4.5 W·m-2, 升温幅度将达1.1~2.6 ℃, 在RCP6.0情景下2100年辐射强迫达6.0 W·m-2, 升温幅度达1.4~3.1 ℃(Taylor et al., 2012; Meinshausen et al., 2011; Warszawaski et al., 2013; Hempel et al., 2013).这些气候变化情景都是到2100年全球大气中辐射强迫分别为2.6、4.5、6.0和8.5 W·m-2, 但在不同区域不同时间段温度和辐射强迫值变化不同, 以及在RCP6.0情景下温度和辐射变化不一定比RCP4.5情景下高.本文研究考虑到了2050年, 没有分析到2100年, 并且本文研究范围为武夷山自然保护区, 与全球2100年各个辐射强迫下温度变化和辐射强度变化不一定相同.因此, 在分析未来气候变化对毛竹异戊二烯日和年排放速率影响中, 毛竹异戊二烯日或年排放速率变化幅度在RCP6.0情景下不一定比RCP4.5情景下低.另外, 在未来气候变化对毛竹异戊二烯日排放速率影响中, 由于同时受温度变化和辐射强度影响, 在各个情景下温度和辐射存在波动, 所以也导致毛竹异戊二烯日或年排放速率变化幅度在RCP6.0情景下不一定比RCP4.5情景下高, 但在RCP8.5情景下毛竹异戊二烯日排放速率却最高.这反映了未来气候变化在全球、区域与地区尺度及不同时间段各个要素变化程度不同.
本研究结果反映气候变化影响下毛竹异戊二烯排放速率增加, 意味着大气中VOCs浓度的变化及对应的大气化学过程速率改变和大气环境的改变.特别是气候变化下异戊二烯排放速率增加将可能使大气中O3浓度改变, 进而影响空气质量(Possell et al., 2011; Monson et al., 2001), 并且对气候变暖也将造成反馈(Collins et al., 2002; Millet et al., 2016).这需要在预测未来大气环境变化和计算植被BVOCs清单中需要考虑.当然, 也有研究发现, 气候变化对BVOCs排放的影响不确定.如Schollert等(2015)研究发现, 气候变化改变了极地植物解剖结构, 但对植物BVOCs排放却没有影响,说明植物发育过程影响了BVOCs排放, 对此需要在以后的研究中进一步分析.
一些研究认为, 气候变化下植物生物量增加导致异戊二烯排放增加(Constable et al., 1999).本研究没有考虑气候变化下生物量的改变, 如果考虑气候变化对生物量的影响, 可以推断气候变化对毛竹异戊二烯排放的影响更大.另外, 气候变化下植物异戊二烯排放速率的改变与植物生理活动有关.如研究发现, 植物异戊二烯排放与光合作用关系密切, 在植物光合作用过程中当碳不受限制时, 异戊二烯排放与ATP含量高度相关(Loreto et al., 1993), 大气中CO2浓度增加将抑制植物异戊二烯排放, 抵消由于地上生物量增加而增加的异戊二烯排放量(Monson et al., 2007);在高温和CO2下植物VOCs排放与光合作用和叶绿素结构关系密切(Velikova et al., 2009).另外, 气候变化下植物异戊二烯排放速率变化与植物解剖结构也有密切关系.如有研究发现, 温度升高和O3浓度变化下叶片结构与植物BVOCs排放关系密切(Hartikainen et al., 2009), 并且植物地下结构与植物BVOCs排放也有密切关系(Henke et al., 2015).还有研究发现, 气候变化引起植物解剖结构改变, 但对BVOCs排放并没有影响(Schollert et al., 2015).也有研究发现, 变暖和增加积雪使光合作用减少, 但对BVOCs排放却没有影响, 主要是因为改变了叶片结构(Schollert et al., 2017).气候变化下植物叶片发育不同也将导致VOCs排放不同(Brilli et al., 2016).本研究中, 气候变化对毛竹异戊二烯排放的影响可能与气候变化下毛竹叶片生理活动与解剖结构变化有关, 这种影响需要在以后的研究中认真考虑.
一些研究发现, 气候变化引起的干旱对植物BVOCs排放影响较大(Faubert et al., 2010), 水分改变对植物VOCs排放影响明显(Filella et al., 2009), 干旱会导致植物BVOCs排放增加(Seco et al., 2015;Geron et al., 2016).另外, 一些研究发现, CO2浓度变化(Potosnak et al., 2014; Possell et al., 2011)及温度和CO2浓度变化(Potosnak et al., 2014)或O3和CO2浓度变化(Calfapietra et al., 2007)对植物异戊二烯排放也有一定的影响, 并且土地利用、CO2浓度、植被组成和气候变化对异戊二烯排放有综合的影响(Arneth et al., 2008).本研究中主要考虑了温度和辐射强度变化对毛竹异戊二烯排放的影响, 没有考虑其它因素变化对毛竹异戊二烯排放的影响.同时, 未来气候变化情景下云的变化对有效辐射进而对植被异戊二烯排放的影响, 以及大气气溶胶潜在变化对光合有效辐射及植被异戊二烯排放的影响等都需要进行深入研究.
气象因素误差可能对植物异戊二烯排放模拟结果造成一定的影响(司徒淑娉等, 2010).本文在模拟植物异戊二烯排放中, 主要考虑了温度和光合有效辐射量子密度.植物叶片温度与气温有关, 但存在一定差异.在本研究中, 由于缺乏叶温观测资料, 而利用能量守衡方法计算叶温较复杂, 因而参考文献中的做法(张莉等, 2003; 周秀骥等, 2004), 忽略了叶片温度和气温差异, 这可能对研究结果造成一定误差.需要说明的是, 要精确估算植物BVOCs排放, 理想情况是了解每个叶片温度.由于辐射差异造成的能量平衡不同, 气温和叶温在不同时间差异不同, 在白天气温比叶温低, 而夜间却是叶温比气温低(周秀骥等, 2004).另外, 叶气温差是叶片特征尺度、气孔导度、饱和水气压差和净辐射等的函数.无论气孔导度有多大, 叶片越小, 其温度与气温相差越小;当气孔关闭时, 大叶片的叶温比小叶片更高;当气孔开放时, 大叶片的叶温比小叶片更低; 对宽大的叶片, 当气孔开放时, 叶温和气温趋于相等, 而与叶片大小无关(胡继超等, 2014).从宏观上分析气候变化对毛竹异戊二烯排放的影响, 这些误差是可以忽略的.此外, 植物光合有效辐射强度与太阳辐射强度密切相关, 但在植物冠层中辐射强度并不是均匀的, 处在冠层不同位置的叶片接受到的光合有效辐射量子密度可能不同.在本文中, 计算毛竹叶片中光合有效辐射量子密度时没有区分冠层不同位置叶片的差异, 而是按照平均值计算, 这也可能对研究结果造成一定的误差.不过, 从分析气候变化对植物异戊二烯排放影响的宏观趋势来看, 这些误差也可以忽略.
需要指出的是, 本研究只采用了单一气候模式模拟的气候变化情景结果, 不同模式下结果可能存在一定的差异, 需要在以后的研究中进一步分析.另外, 一些研究发现, 气候变化会引起植被变化导致BVOCs排放变化(Valolahti et al., 2015), 或气候变化会改变植物组成使BVOCs排放改变(Faubert et al., 2012; Vedel-Petersen et al., 2012).而本研究只考虑了在武夷山自然保护区分布的毛竹, 没有考虑其他区域毛竹异戊二烯的排放, 并且本研究也没有考虑气候变化可能导致毛竹分布范围改变而对异戊二烯排放造成的影响.这样也可能存在一定的误差, 需要在以后的研究中考虑这些问题.尽管如此, 本研究利用新的气候变化情景数据, 分析了未来气候变化对毛竹异戊二烯排放速率的影响, 对科学认识气候变化对植物BVOCs排放及大气环境影响仍有一定的参考意义.
5 结论(Conclusions)1) 在未来气候变化下, 武夷山自然保护区年均气温呈上升趋势, 年降水量和辐射强度波动较大, 呈增长或下降趋势.
2) 气候变化下, 毛竹异戊二烯日排放速率平均值在未来气候变化情景下比基准情景下高约30 μg·g-1·d-1, 在RCP8.5情景下高约48 μg·g-1·d-1;毛竹异戊二烯日排放速率在未来气候变化情景与基准情景下的差异在1~90 d和301~365 d较小, 在91~300 d较大;相比基准情景, 未来气候变化情景下毛竹异戊二烯日排放速率在1~190 d和271~365 d增幅较大, 在191~270 d增幅较小, 在RCP8.5情景下增幅最大.
3) 气候变化影响下, 毛竹异戊二烯年排放速率在未来气候变化情景下比基准情景下约高10000 μg·g-1·a-1以上, 在RCP8.5情景下比基准情景下约高13%.
Arneth A, Schurgers G, Hickler T, et al. 2008. Effects of species composition, land surface cover, CO2concentration and climate on isoprene emissions from European forests[J]. Plant Biology, 10(1): 150–162.
DOI:10.1055/s-2007-965247
|
白建辉, TiffanyD, 余树全, 等. 2015. 亚热带竹林挥发性有机物排放的模拟[J]. 生态环境学报, 2015, 24(12): 1923–1937.
|
白建辉, 林凤友, 万晓伟, 等. 2012. 长白山温带森林挥发性有机物的排放通量[J]. 环境科学学报, 2012, 32(3): 545–554.
|
Brilli F, Gioli B, Fares S, et al. 2016. Rapid leaf development drives the seasonal pattern of volatile organic compound (VOC) fluxes in a 'coppiced' bioenergy poplar plantation[J]. Plant, Cell & Environment, 39(3): 539–555.
|
Calfapietra C, Wiberley A E, Falbel T G, et al. 2007. Isoprene synthase expression and protein levels are reduced under elevated O3 but not under elevated CO2(FACE) in field-grown aspen trees[J]. Plant, Cell & Environment, 30(5): 654–661.
|
陈颖, 李德文, 史奕, 等. 2009. 沈阳地区典型绿化树种生物源挥发性有机物的排放速率[J]. 东北林业大学学报, 2009, 37(3): 47–49.
|
Collins W J, Derwent R C, Johnson C E, et al. 2002. The oxidation of organic compounds in the troposphere and their global warming potentials[J]. Climatic Change, 52: 453–479.
DOI:10.1023/A:1014221225434
|
Constable J V H, Guenther A B, Schimel D S, et al. 1999. Modelling changes in VOC emission in response to climate change in the continental United States[J]. Global Change Biology, 5(7): 791–806.
DOI:10.1046/j.1365-2486.1999.00273.x
|
Copolovici L, Niinemets V. 2010. Flooding induced emissions of volatile signalling compounds in three tree species with differing water logging tolerance[J]. Plant, Cell & Environment, 33(9): 1582–1594.
|
Ephrath J E, Goudriaan J, Marani A. 1996. Modelling diurnal patterns of air temperature, radiation wind speed and relative humidity by equations from daily characteristics[J]. Agricultural Systems, 51(4): 377–393.
DOI:10.1016/0308-521X(95)00068-G
|
Faubert P, Tiiva P, Rinnan Å, et al. 2010. Doubled volatile organic compound emissions from subarctic tundra under simulated climate warming[J]. New Phytologist, 187(1): 199–208.
DOI:10.1111/j.1469-8137.2010.03270.x
|
Faubert P, Tiiva P, Michelsen A, et al. 2012. The shift in plant species composition in a subarctic mountain birch forest floor due to climate change would modify the biogenic volatile organic compound emission profile[J]. Plant and Soil, 352(1/2): 199–215.
|
Fehsenfeld F, Callvert J, Fall R, et al. 1992. Emissions of volatile of organic compounds from vegeration and the implications for atmospheric chemistry[J]. Global Biochemical Cycles, 6(4): 389–430.
DOI:10.1029/92GB02125
|
Filella I, Peñuelas J, Seco R. 2009. Short-chained oxygenated VOC emissions in Pinus halepensis in response to changes in water availability[J]. Acta Physiologiae Plantarum, 31(2): 311–318.
DOI:10.1007/s11738-008-0235-6
|
Filella I, Wilkinson M J, Llusià J, et al. 2007. Volatile organic compounds emissions in Norway spruce (Picea abies) in response to temperature changes[J]. Physiologia Plantarum, 130(1): 58–66.
DOI:10.1111/ppl.2007.130.issue-1
|
Geron C, Daly R, Harley P, et al. 2016. Large drought-induced variations in oak leaf volatile organic compound emissions during PINOT NOIR[J]. Chemosphere, 146: 8–21.
DOI:10.1016/j.chemosphere.2015.11.086
|
Guenther A B, Zimmerman P R, Harley P C, et al. 1993. Isoprene and monoterpene emission rate variability:model evaluations and sensitivity analyses[J]. Journal of Geophysical Research, 98(D7): 12609–12617.
DOI:10.1029/93JD00527
|
Guenther A, Hewitt C N, Erickson D, et al. 1995. A global model of natural volatile organic compounds emissions[J]. Journal of Geophysical Research, 100: 8873–8892.
DOI:10.1029/94JD02950
|
Harley P, Vasconcellos P, Vierling L, et al. 2004. Variation in potential for isoprene emissions among neotropical forest sites[J]. Global Change Biology, 10: 630–650.
DOI:10.1111/gcb.2004.10.issue-5
|
Hartikainen K, Riikonen J, Nerg A M, et al. 2012. Impact of elevated temperature and ozone on the emission of volatile organic compounds and gas exchange of silver birch (Betula pendula Roth)[J]. Environmental and Experimental Botany, 84: 33–43.
DOI:10.1016/j.envexpbot.2012.04.014
|
Hartikainen K, Nerg A M, Kivimaenpaa M, et al. 2009. Emissions of volatile organic compounds and leaf structural characteristics of European aspen (Populus tremula) grown under elevated ozone and temperature[J]. Tree Physiology, 29(9): 1163–1173.
DOI:10.1093/treephys/tpp033
|
Hempel S, Frieler K, Warszawski L, et al. 2013. A trend-preserving bias correction-the isi-mip approach[J]. Earth System Dynamoical, 42920: 219–236.
|
Henke A, Kunert M, Veit D, et al. 2015. Analysis of volatiles from Picea abies triggered by below-ground interactions[J]. Environmental and Experimental Botany, 110: 56–61.
DOI:10.1016/j.envexpbot.2014.09.009
|
胡继超, 申双和, 孙卫国, 等. 2014. 微气象学基础[M]. 北京: 气象出版社: 216–217.
|
Kellomäki S, Rouvinen I, Peltola H, et al. 2001. Impact of global warming on the tree species composition of boreal forests in Finland and effects on emissions of isoprenoids[J]. Global Change Biology, 7(5): 531–544.
DOI:10.1046/j.1365-2486.2001.00414.x
|
Kramshøj M, Vedel-Petersen I, Schollert M, et al. 2016. Large increases in Arctic biogenic volatile emissions are a direct effect of warming[J]. Nature Geoscience, 9(5): 349–352.
DOI:10.1038/ngeo2692
|
Laffineur Q, Aubinet M, Schoon N, et al. 2013. Impact of diffuse light on isoprene and monoterpene emissions from a mixed temperate forest[J]. Atmospheric Environment, 74: 385–392.
DOI:10.1016/j.atmosenv.2013.04.025
|
Laothawornkitkul J, Taylor J E, Paul N D, et al. 2009. Biogenic volatile organic compounds in the earth system[J]. New Phytologist, 183(1): 27–51.
DOI:10.1111/nph.2009.183.issue-1
|
Lathière J, Hauglustaine D A, Noblet-DucoudréDe N, et al. 2005. Past and future changes in biogenic volatile organic compound emissions simulated with a global dynamic vegetation model[J]. Geophysical Research Letters.
DOI:10.1029/2005GL024164
|
Levis S, Wiedinmyer C, Bonan G B, et al. 2003. Simulating biogenic volatile organic compound emissions in the community climate system model[J]. Journal of Geophysical Research, 108: 4659.
DOI:10.1029/2002JD003203,D21
|
Lindwall F, Schollert M, Michelsen A, et al. 2016. Fourfold higher tundra volatile emissions due to arctic summer warming[J]. Journal of Geophysical Research:Biogeosciences, 121(3): 895–902.
DOI:10.1002/2015JG003295
|
Lindwall F, Svendsen S S, Nielsen C S, et al. 2016. Warming increases isoprene emissions from an arctic fen[J]. Science of the Total Environment, 553: 297–304.
DOI:10.1016/j.scitotenv.2016.02.111
|
Loreto F, Sharkey T D. 1993. On the relationship between isoprene emission and photosynthetic metabolites under different environment conditions[J]. Planta, 189: 420–424.
DOI:10.1007/BF00194440
|
梁宝生, BaraldiR, RappariniF. 2003. 夏季黄连木排放萜烯类化合物浓度日变化及排放速率研究[J]. 北京大学学报, 2003, 39(4): 517–521.
|
梁珍海, 刘海燕, 陈霞, 等. 2011. 南京紫金山不同植物群落VOCs组成[J]. 南京林业大学学报, 2011, 25(1): 4–38.
|
Meinshausen M, Smith S J, Calvin K, et al. 2011. The RCP greenhouse gas concentrations and their extensions from 1765 to 2300[J]. Climate Change, 109(1/2): 213–241.
|
Millet D B, Baasandorj M, Hu L, et al. 2016. Nighttime chemistry and morning isoprene can drive urban ozone downwind of a major deciduous forest[J]. Environmental Science & Technology, 50(8): 4335–4342.
|
Monson R K, Trahan N, Rosenstiel T N, et al. 2007. Isoprene emission from terrestrial ecosystems in response to global change:minding the gap between models and observations[J]. Philosophical Transactions of the Royal Society A:Mathematical, Physical and Engineering Sciences, 365: 1677–1695.
DOI:10.1098/rsta.2007.2038
|
Monson R K, Holland E A. 2001. Biospheric trace gas fluxes and their control over tropospheric chemistry[J]. Annual Review of Ecology and Systematics, 32: 547–576.
DOI:10.1146/annurev.ecolsys.32.081501.114136
|
Monson R K, Grote R, Niinemets V, et al. 2012. Modeling the isoprene emission rate from leaves[J]. New Phytologist, 195: 541–559.
DOI:10.1111/j.1469-8137.2012.04204.x
|
宁平, 郭霞, 田森林, 等. 2013. 昆明地区典型乔木主要挥发性有机物释放规律[J]. 中南大学学报, 2013, 44(3): 1290–1296.
|
Pegoraro E, Rey A, Abrell L, et al. 2006. Drought effect on isoprene production and consumption in biosphere 2 tropical rainforest[J]. Global Change Biology, 12: 456–469.
DOI:10.1111/gcb.2006.12.issue-3
|
Peñuelas J, Filella I, Seco R, et al. 2009. Increase in isoprene and monoterpene emissions after re-watering of droughted Quercus ilex seedlings[J]. Biologia Plantarum, 53(2): 351–354.
DOI:10.1007/s10535-009-0065-4
|
Possell M, Hewitt C N. 2011. Isoprene emissions from plants are mediated by atmospheric CO2 concentrations[J]. Global Change Biology, 17: 1595–1610.
DOI:10.1111/gcb.2011.17.issue-4
|
Potosnak M J, Stourgeon L L, Nunez O. 2014. Increasing leaf temperature reduces the suppression of isoprene emission by elevated CO2 concentration[J]. Science of the Total Environment, 481: 352–359.
DOI:10.1016/j.scitotenv.2014.02.065
|
Potter C S, Alexander S E, Coughlan J C, et al. 2001. Modeling biogenic emissions of isoprene:exploration of model drivers, climate control algorithms, and use of global satellite observations[J]. Atmospheric Environment, 35: 6151–6165.
DOI:10.1016/S1352-2310(01)00390-9
|
Rinnan R, Rinnan Å, Faubert P, et al. 2011. Few long-term effects of simulated climate change on volatile organic compound emissions and leaf chemistry of three subarctic dwarf shrubs[J]. Environmental and Experimental Botany, 72: 377–386.
DOI:10.1016/j.envexpbot.2010.11.006
|
Schollert M, Kivimäenpää M, Valolahti H M, et al. 2015. Climate change alters leaf an atomy, but has no effects on volatile emissions from arctic plants[J]. Plant, Cell & Environment, 38: 2048–2060.
|
Schollert M, Kivimäenpää M, Michelsen A, et al. 2017. Leaf anatomy, BVOC emission and CO2 exchange of arctic plants following snow addition and summer warming[J]. Ann Bot, 119(3): 433–445.
DOI:10.1093/aob/mcw237
|
Seco R, Karl T, Guenther A, et al. 2015. Ecosystem-scale volatile organic compound fluxes during an extreme drought in a broadleaf temperate forest of the Missouri Ozarks (central USA)[J]. Global Change Biology, 21: 3657–3674.
DOI:10.1111/gcb.12980
|
盛裴轩, 毛节泰, 李建国, 等. 2003. 大气物理学[M]. 北京: 北京大学出版社.
|
司徒淑娉, 王雪梅, GuentherA, 等. 2009. 典型夏季珠江三角洲地区植被的异戊二烯排放[J]. 环境科学学报, 2009, 29(4): 822–829.
|
司徒淑娉, 王雪梅, GuentherA, 等. 2010. 气象模拟误差对异戊二烯排放估算的影响[J]. 环境科学学报, 2010, 30(12): 2383–2391.
|
Stocker T F, Qin D, Plattner G K. 2013. Climate Change 2013:The Physical Science Basis. Contribution of Working Group Ⅰ to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change[M]. Cambridge, UK and New York, NY, USA: Cambridge University Press: 4–23.
|
Taylor K E, Stouffer R J, Meehl G A. 2012. An overview of CMIP5 and experiment design[J]. Bulletin of the American Meterological Society, 93(40): 485–498.
|
唐孝炎, 张远航, 邵敏, 等. 2010. 大气环境化学(第2版)[M]. 北京: 高等教育出版社.
|
Tiiva P, Faubert P, Michelsen A, et al. 2008. Climatic warming increases isoprene emission from a subarctic heath[J]. New Phytologist, 180(4): 853–863.
DOI:10.1111/nph.2008.180.issue-4
|
Turner D P, Baglio J V, Wones A, et al. 1991. Climate change and isoprene emissions from vegetation[J]. Chemosphere, 23(1): 37–56.
DOI:10.1016/0045-6535(91)90115-T
|
Unger N. 2013. Isoprene emission variability through the twentieth century[J]. Journal of Geophysical Research:Atmospheres, 118(13): 606–13613.
|
Valolahti H, Kivimäenpää M, Faubert P, et al. 2015. Climate change-induced vegetation change as a driver of increased subarctic biogenic volatile organic compound emissions[J]. Global Change Biology, 21(9): 3478–3488.
DOI:10.1111/gcb.2015.21.issue-9
|
Vedel-Petersen I, Schollert M, Nymand J, et al. 2015. Volatile organic compound emission profiles of four common arctic plants[J]. Atmospheric Environment, 120: 117–126.
DOI:10.1016/j.atmosenv.2015.08.082
|
Velikova V, Tsonev T, Barta C, et al. 2009. BVOC emissions, photosynthetic characteristics and changes in chloroplast ultrastructure of Platanus orientalis L. exposed to elevated CO2 and high temperature[J]. Environmental Pollution, 157(10): 2629–2666.
DOI:10.1016/j.envpol.2009.05.007
|
Warszawaski L, Frieler K, Huber V, et al. 2013. the inter-sectoral impact model intercomparison project (isi-mip):project framework[J]. Proceedings of the National Academy of Sciences, 119(9): 3228–3232.
|
王效科, 牟玉静, 欧阳志云, 等. 2002. 太湖流域主要植物异戊二烯排放研究[J]. 植物学通报, 2002, 19(2): 224–230.
|
王志辉, 白郁华, 王雪松, 等. 2003. 西双版纳地区三叶橡胶树异戊二烯和单萜烯排放机理初步研究[J]. 北京大学学报, 2003, 39(4): 512–516.
|
王志辉, 张树宇, 陆思华, 等. 2003. 北京地区植物VOCs排放速率的测定[J]. 环境科学, 2003, 24(2): 7–12.
|
吴建国. 2017. 气候变化对生物多样性影响与风险研究[M]. 北京: 科学出版社.
|
闫雁, 王志辉, 白郁华, 等. 2005. 中国植被VOC排放清单的建立[J]. 中国环境科学, 2005, 25(1): 110–114.
|
Zhang Y, Hu X M, Leung L R, et al. 2008. Impacts of regional climate change on biogenic emissions and air quality[J]. Journal of Geophysical Research:Atmospheres (1984-2012), 113: D18.
|
张莉, 王效科, 欧阳志云, 等. 2003. 中国森林生态系统的异戊二烯排放研究[J]. 环境科学, 2003, 24(1): 8–15.
|
周秀骥, 唐孝炎, 冯宗炜, 等. 2004. 长江三角洲低层大气与生态系统相互作用研究[M]. 北京: 气象出版社.
|
周允华, 项月琴, 栾禄凯. 1996. 光合有效量子密度气候学计算[J]. 气象学报, 1996, 54(4): 447–455.
|