 2018, Vol. 38
2018, Vol. 38



2. 陕西秦岭森林生态系统国家野外科学观测研究站, 安康 711600;
3. 西北农林科技大学林学院生态预测与全球变化实验室, 杨凌 712100;
4. 中国科学院生态环境研究中心, 北京 100085
2. Qinling National Forest Ecosystem Research Station, Ankang 711600;
3. Laboratory for Ecological Forecasting and Global Change, College of Forestry, Northwest A & F University, Yangling 712100;
4. Research Center for Eco-environmental Sciences, Chinese Academy of Sciences, Beijing 100085
近年来, 大气中CO2、CH4和N2O等温室气体的浓度加速升高, 由此引发的温室效应已成为人们普遍关注的环境问题(Lal, 2004).森林作为陆地生态系统的主体, 对大气环境起着至关重要的调控作用, 森林土壤更是温室气体重要的源或汇(West et al., 2002).研究表明, CO2通量与土壤温度、土壤水分正相关(Wang et al., 2003; Davidson et al., 2006; Rosenkranz et al., 2006), CH4和N2O通量主要取决于土壤微生物的生命活动进程(Ullah et al., 2015).温带森林中, 地形、水分等环境因子的综合作用会导致CO2、CH4和N2O通量出现较大的时空变异(Mcclain et al., 2003), 尤其是地形的变化会引起土壤温度、水分、pH和元素含量等因子产生差异, 从而在局部范围内形成若干个微环境, 最终导致土壤CO2、CH4、N2O通量因地形不同而呈现不同的时空变异.Hirobe等(1998)发现, 不同坡位土壤微生物的净硝化速率受控于土壤水分, 而土壤水分受到地形因素的影响, 上坡位的土壤排水性优于下坡位, 下坡位较为湿润的土壤环境更有利于硝化细菌的生长发育, 因而下坡位处N2O排放量大于上坡位(Corre et al., 2002; Osaka et al., 2015; Mcswiney et al., 2001).Fang等(2009)推测, 由于有效水和营养物质含量的不同, 下坡位土壤的CO2、N2O排放量大于上坡位, CH4吸收量则小于上坡位, 然而在Fang等的实验中, 只有N2O遵循这一规律.Peichl等(2010)在加拿大温带针叶林的研究发现, 土壤CO2通量与地温正相关, CH4、N2O通量未表现出与地温的相关性.
坡位要素对土壤温室气体通量产生影响的途径通常具有不确定性, 在上述研究中(Hirobea et al., 1998; Corre et al., 2002; Osaka et al., 2015; Mcswiney et al., 2001; Fang et al., 2009; Peichl et al., 2010), 土壤温室气体通量的差异主要源于不同坡位条件下环境因子(土壤水分、营养物含量和地温)的不同.秦岭作为我国南北气候的分界线和长江、黄河两大水系的分水岭, 对秦岭森林生态系统的研究在全国森林生态系统总体研究中占据着重要地位.但目前关于秦岭地区坡位要素对土壤温室气体通量影响的研究还未见报道.在地形起伏变化较大的地区, 尤其是山区, 根据地形划分不同的小单元(上坡位、中坡位和下坡位)来研究CO2、CH4、N2O通量及其影响因素, 将能更加准确地估算整个地区的CO2、CH4、N2O通量和深入理解地形对其影响的机理.本研究在秦岭火地塘林区选取分布较广的天然次生油松林, 对林内上、中、下3个坡位土壤主要温室气体通量进行为期1年的观测, 旨在研究秦岭林区土壤CO2、CH4、N2O通量在不同坡位的季节变化特征及其与环境因子的相互关系.研究结果将有利于更加准确地估算秦岭地区森林土壤CO2、CH4、N2O通量, 使我国(京都议定书缔约国)在应对气候变化谈判中处于更加有利的地位.
2 材料与方法(Materials and methods) 2.1 研究区概况秦岭火地塘林区地处秦岭南坡中段(33°18′~33°28′ N, 108°21′~108°39′ E), 属暖温带与北亚热带的过渡地带, 森林植被类型主要为寒温带针叶林和温带针阔混交林, 森林覆盖率91.8%, 海拔800~2500 m, 年日照时间约为1100~1300 h, 年均气温9 ℃, 无霜期170 d, 林区土壤类型有山地草甸土、暗棕壤和山地棕壤(李玉田等, 2017).
试验地位于秦岭火地塘林区的天然次生油松林内, 海拔1585 m, 年降水量939 mm, 年平均气温8~10 ℃, 主要树种有油松(Pinus tabulaeformis)、华山松(Pinus armandii)、锐齿栎(Quercus aliena var. acuteserrata).样地内分别设上坡位(西南坡, 40°)、中坡位(西南坡, 36°~38°)、下坡位(西坡, 30°)3个样方.
2.2 研究方法本研究在秦岭火地塘林区天然次生油松林内选取上、中、下3个坡位, 每个坡位设置一个20 m×20 m的样方, 在样方内选取4个重复, 采用静态箱-气相色谱法对土壤温室气体(CO2、N2O、CH4)通量进行1年的监测(图 1).监测期内共采集气样20次, 其中, 植物生长季(2015年4—9月)每月采气2次, 非生长季(2014年10月—2015年3月)每月采气1次, 冻融交替期每周采气1次.采气箱高度为35 cm, 底面积为50 cm×50 cm, 内芯为不锈钢板, 厚度为2 mm, 外覆隔热材料, 箱盖上装有采气三通阀、采气用的胶皮管和气压平衡管(内径4 mm, 长4 cm)(Hutchinson, 2004).采气步骤如下:实验开始前两周将不锈钢底座插入样地土壤, 每个样地放置4个底座, 采样时将采样箱扣入底座水槽内, 轻微挪动使采样箱边缘与底座凹槽彻底吻合, 注水密封.在采样箱正上方悬挂温度计, 用于记录采样前、后的气温, 在采气箱密封后的第0、10、20、30 min, 打开平衡管, 用手持式采气泵抽取采样箱内气体于500 mL的密封气袋中, 每次采气结束后关闭平衡管(庞军柱等, 2010).所采气样在1周内带回实验室用气相色谱仪进行分析(Agilent Technologies公司生产的Agilent7890AGC System气相色谱仪).采样点的土壤水分(0~20 cm、20~40 cm)和土壤温度(0~10 cm)分别用烘干法和SN2022型数字温度计测定(读数分辨率:0.1 ℃, 测量准确度:0.5 ℃, 温度范围:-30~50 ℃).试验地土壤容重两月一测, 并根据公式计算土壤孔隙充水率.不同土层的(0~20 cm、20~40 cm)硝态氮(NO3-)和铵态氮(NH4+)含量用连续流动分析仪测定(SANT++, Skalar Company, the Netherlands).气温和降雨数据由陕西秦岭森林生态系统国家野外科学观测研究站提供.
 |
| 图 1 不同坡位示意图 Fig. 1 Schematic drawing of the designated slope position along the study slope |
| 表 1 各坡位土壤理化性质 Table 1 Soil characteristics at different slope positions |
温室气体通量的计算公式如下:
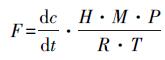
|
(1) |
式中, F表示CO2(mg · m-2 · h-1)、CH4(μg · m-2 · h-1)、N2O(μg · m-2 · h-1)的通量值, dc/dt表示某一时段内采样箱中温室气体的浓度动态(nL · L-1 · h-1), H为采样箱高度(m), M为CO2、CH4、N2O的摩尔质量(g · mol-1), P为大气压(MPa), R为气体常数(cm3 · MPa · K-1 · mol-1), T为通量箱内气温(K).
温室气体累积量Xn计算公式如下:
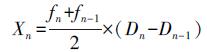
|
(2) |
式中, Xn表示试验地温室气体在某一时间范围内的总累积量(CO2单位为mg · m-2, CH4、N2O单位为μg · m-2);fn-1表示该时段第1天所监测的试验地温室气体通量, fn表示该时段最后1天所监测的试验地温室气体通量, 二者的平均数表示该时段试验地温室气体的平均通量, CO2单位为mg · m-2 · d-1, CH4、N2O单位为μg · m-2 · d-1;Dn-1表示该间段第1天, Dn表示时段的最后1天, Dn-Dn-1表示该时段的总天数.
土壤孔隙充水率(WFPS)计算公式(Merino et al., 2004)为:
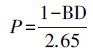
|
(3) |
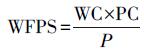
|
(4) |
式中, P表示土壤总孔隙度, WC表示土壤质量含水率, BD表示土壤容重(g · cm-3), 2.65表示土壤密度(g · cm-3).
全球增温潜势(GWP, t · hm-2)计算公式(Peichl et al., 2010)为:
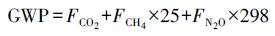
|
(5) |
式中, GWP表示全球增温潜势值, FCO2表示CO2年通量(t · hm-2 · a-1), FCH4表示CH4年通量(kg · hm-2 · a-1), FN2O表示N2O年通量(kg · hm-2 · a-1).
2.4 统计分析采用SPSS 21.0进行数据统计分析, 利用Pearson相关分析研究温室气体通量与环境要素之间的关系, 利用Origin 2016进行作图, 利用DPS数据处理系统对不同坡位间的温室气体通量进行one-way ANOVA比较.
3 结果(Results) 3.1 油松林不同坡位的环境因子动态秦岭火地塘林区2014年10月—2015年9月降水量939 mm, 6、7月累计降水312 mm, 占全年降水的1/3, 年均气温8~10 ℃, 雨热同期(图 2);土壤温度(0~10 cm)受气温的影响较大, 具有明显的季节性规律, 最高温度为17.2 ℃(2015年6月27日), 最低温度为0.05 ℃(2015年2月28日), 3个坡位土壤温度变化基本一致(图 3).
 |
| 图 2 油松林样地降雨和日气温 Fig. 2 Maximum and minimum daily air temperature and daily precipitation at the study site |
 |
| 图 3 油松林样地不同坡位地温的变化(0~10 cm) Fig. 3 Soil temperature of the study sites(0~10 cm) |
上、下坡位0~20 cm土壤孔隙充水率(WFPS)长期维持在30%~60%, 没有表现出明显的季节性规律;而中坡位(0~20 cm)WFPS维持在60%~95%, 并在非生长季达到最大值95%(图 4a), 远高于上、下坡位的WFPS, 这主要是因为:①中坡位植被覆盖度高、腐殖质层厚, 林下土壤接收的太阳辐射小于上、下坡位, 故中坡位处土壤水分的蒸发量相对较小;②中坡位土壤粒径最小, 粒径小的土壤排水性更差, 土壤的持水能力相对较强.以上两点导致中坡位WFPS(60%~95%)高于上、下坡位(30%~60%).各坡位土壤20~40 cm处WFPS均在30%~70%范围内波动, 值得注意的是, 5月1日试验地降雨34.1 mm, 上坡位WFPS因此升高至65%左右, 并随着降雨事件一直持续至6月初.下、中坡位WFPS虽在生长季内分别达到全年最大值57%~69%, 但整体来看, 并未随季节发生有规律的变化(图 4b).
 |
| 图 4 土壤孔隙充水率的变化(a.0~20 cm, b.20~40 cm) Fig. 4 Soil water filled pore space of the study sites(a.0~20 cm, b.20~40 cm) |
油松林表层土壤(0~20 cm)的硝态氮含量具有明显的季节性规律, 进入生长季后, 土壤硝态氮含量快速升高且波动较大, 尤其是上坡位土壤硝态氮含量在5月出现连续上涨, 6月7日回落到以前的水平, 这一现象与同时期的WFPS变化相对应:2015年5月1日, 试验地降雨34.2 mm, 在2015年5月3日的监测中, 上坡位WFPS由26.63%跃升至59.10%, 土壤水分的上升促使硝化细菌活性增强, 土壤硝态氮含量由0.09 μg · g-1上升至34.03 μg · g-1;2015年5月21—22日, 试验地累计降雨25.5 mm, 次日上坡位WFPS为45.67%, 该范围的土壤水分含量一方面有利于硝化作用的顺利进行, 同时又不会因土壤水分含量过高而阻碍了O2在土壤孔隙间的传输, 为硝化细菌提供了理想的水氧环境, 因而土壤硝态氮含量在该日达到最大值58.82 μg · g-1;2015年6月7日, 上坡位WFPS回落至27%, 土壤硝态氮含量随之下降为1.79 μg · g-1.油松林土壤铵态氮含量在非生长季内波动较大, 生长季内虽出现多个小高峰, 但总体变动不大(图 5b).各坡位土壤硝态氮含量的年平均值大小依次为:中坡位(18.06 μg · g-1)>下坡位(9.23 μg · g-1)>上坡位(6.43 μg · g-1);铵态氮含量的年平均值大小依次为:下坡位(24.81 μg · g-1)>中坡位(16.52 μg · g-1)>上坡位(13.67 μg · g-1)(图 5b).
 |
| 图 5 表层土壤(0~20cm)硝态氮(a)、铵态氮(b)含量 Fig. 5 Soil nitrate content (a) and soil ammonium content (b)(0~20 cm)of the study sites |
从全年来看, 油松林土壤CO2通量表现为倒“S”形变化, 具有明显的季节变化规律(图 6a).进入生长季, 油松林土壤理想的水热条件使得土壤微生物与植物根系的生命活动逐步增强, 在该过程中伴随着CO2的大量排放.2015年8月12日, 下坡位、中坡位同时达到CO2排放最大值, 分别为349.35、309.85 mg · m-2 · h-1;上坡位则于2015年6月26日达到最大值333.61 mg · m-2 · h-1.生长季末, 低温不利于土壤微生物与植物根系的生命活动, 从而出现CO2排放量减少的现象, 这一过程在温带森林中极为普遍.非生长季, CO2排放量降至最低, 下坡位于2015年3月8日出现CO2排放最小值33.94 mg · m-2 · h-1;2014年12月27日, 中坡位出现最小值29.26 mg · m-2 · h-1;上坡位则于2015年2月10日出现最小值34.73 mg · m-2 · h-1.各坡位CO2年总排放量从大到小依次为:上坡位(14.17±1.30) t · hm-2、下坡位(13.71±0.85) t · hm-2和中坡位(12.29±1.48) t · hm-2(表 2), 3个坡位之间的土壤CO2通量不存在显著性差异(表 3).
 |
| 图 6 不同坡位油松林土壤CO2 (a)、N2O (b)和CH4 (c)通量的季节动态 Fig. 6 Seasonal patterns of CO2(a), N2O(b) and CH4(c) emissions at different slope positions in Pinus tablaeformis forest |
| 表 2 不同坡位土壤CO2、CH4、N2O年通量及GWP Table 2 Annual acumulative fluxes of CO2, CH4 and N2O at different slope positions and the values of global warming potential |
| 表 3 土壤CO2、CH4、N2O通量在不同坡位间的比较 Table 3 The average fluxes of CO2, CH4, N2O at different slope positions |
不同坡位油松林土壤N2O通量具有一定的季节变化规律.生长季内, N2O通量波动不大, 受冻融循环的影响, N2O在非生长季内排放量较大(图 6b).中坡位土壤最为明显, 2015年1月22日, 中坡位出现N2O排放最大值811.37 μg · m-2 · h-1;2015年9月22—24日试验地累计降雨13.3 mm, 次日(2015年9月25日), 上、下坡位均出现坡位内N2O排放最大值, 分别为45.65、34.37 μg · m-2 · h-1, 值得注意的是, 中坡位土壤N2O最大吸收值130.72 μg · m-2 · h-1同样在该日出现.下坡位的最大吸收值为3.77 μg · m-2 · h-1, 出现在2015年1月22日, 上坡位的最大吸收值为7.57 μg · m-2 · h-1, 出现在2015年3月25日.总体来说, 油松林土壤是N2O的排放源, 各坡位N2O年总排放量依次为中坡位((0.84±0.06) kg · hm-2)>下坡位((0.72±0.09) kg · hm-2)>上坡位((0.69±0.06) kg · hm-2)(表 2), 且土壤N2O通量在中、上坡位间的差异性达到极显著水平(p < 0.01, 表 3).
油松林不同坡位土壤CH4通量未表现出明显的季节变化规律(图 6c).生长季内CH4吸收峰值的出现总是伴随着降雨事件的发生:2015年4月1日, 试验地降雨34.9 mm, 同日下坡位出现CH4最大吸收值428.1 μg · m-2 · h-1;2015年6月6日试验地降雨0.2 mm, 次日CH4吸收值为87.15 μg · m-2 · h-1;2015年8月24日试验地降雨10.1 mm, 次日CH4吸收值为102.13 μg · m-2 · h-1;2015年9月24日试验地降雨9.6 mm, 次日上坡位土壤CH4吸收值为105.35 μg · m-2 · h-1.非生长季, 上、中坡位的CH4最大吸收值均在冻融期内发生:2015年1月22日, 中坡位出现CH4最大吸收值79.64 μg · m-2 · h-1;2014年11月20日上坡位出现CH4最大吸收值142.78 μg · m-2 · h-1, 同日, 中坡位出现CH4最大排放值80.45 μg · m-2 · h-1, 这也是全年唯一一次CH4排放的现象;下坡位、上坡位分别在2015年5月5日、9月12日出现CH4最小吸收值, 分别为30.17、22.9 μg · m-2 · h-1.总体来看, 油松林土壤是CH4的吸收汇, 各坡位CH4年吸收总量依次为下坡位((6.78±1.25) kg · hm-2)>上坡位((4.92±0.59) kg · hm-2)>中坡位((3.84±0.81) kg · hm-2, 表 2).
3.3 不同坡位油松林土壤CO2通量与环境因子的相关关系及全球增温潜势土壤温度、不同土层硝态氮、铵态氮含量及WFPS均会对土壤微生物及植物根系的生命活动造成影响, 从而影响土壤CO2通量, 本研究发现不同坡位油松林土壤CO2通量均与地温显著正相关(p < 0.01, 表 4), 且呈指数型增长(图 7).上坡位土壤CO2通量与WFPS、铵态氮含量(20~40 cm)显著相关(p < 0.05).中坡位CO2通量与硝态氮含量(0~20 cm)、铵态氮含量(20~40 cm)显著相关(p < 0.05)(表 4).不同坡位土壤温室气体的全球增温潜势(GWP)从大到小依次是下坡位>上坡位>中坡位(表 2).
| 表 4 不同坡位土壤CO2、CH4、N2O通量与环境因子的关系 Table 4 The correlation coefficients (r) between CO2, CH4 and N2O fluxes and soil parameters at different slope positions |
 |
| 图 7 油松林土壤CO2通量与土壤温度的相互关系 Fig. 7 The relationship between CO2 emissions and soil temperature in Pinus tablaeformis forest |
本研究中, 各坡位油松林土壤CO2平均通量排序为:上坡位((161.76±14.90) mg · m-2 · h-1)>下坡位((156.49±9.72) mg · m-2 · h-1)>中坡位((140.33±16.90) mg · m-2 · h-1).Xu等(2010)对长白山3种温带森林的土壤CO2通量进行了4年的监测, 发现次生针叶林CO2平均通量为94.83 mg · m-2 · h-1, 所测结果低于本实验数值.这可能是因为相较于长白山地区, 秦岭纬度更低, 森林土壤微生物及植物根系的生命活动相对较强, 故秦岭地区油松林土壤CO2排放量大于长白山地区.试验地土壤CO2排放量与土壤温度显著正相关, 油松林上坡位土壤CO2排放量与WFPS显著相关, 这一结果与其他温带森林的研究结果相一致(Davidson et al., 2002; Savage et al., 1997; Russell et al., 1998).
本研究发现, 土壤质地对各坡位土壤CO2排放有明显影响, 各坡位CO2年排放量大小与土壤粒径排序相一致, 即上坡位(砂壤土)>下坡位(壤土)>中坡位(粉砂壤土), 这是由于疏松的土壤结构有利于促进土壤微生物及植物根系有氧呼吸, 土壤CO2排放量随之增大.油松林上坡位土壤CO2排放量大于中坡位, 与Fang等(2009)的结果相反.这是由于除了土质因素的影响, 还与土壤水分有关, 中坡位WFPS远高于上、下坡位, WFPS过高致使土壤透气性变差(Ryota et al., 2008), 阻碍了土壤孔隙间O2的扩散, 土壤微生物及植物根系因O2供给受限而活性减弱, 土壤CO2排放量随之减小;同时, 土壤微生物及植物根系在土壤内无氧呼吸产生的酒精还会对植物根系造成毒害, 致使植物根系有氧呼吸进一步减弱, 故中坡位土壤的CO2排放量为各坡位间最小值.
4.2 N2O通量在Schmeid等(1987)的研究中, 生长季是N2O的集中排放期, 本研究N2O则是在非生长季大量排放且越冬期占比最大.各坡位的N2O平均通量排序依次为中坡位((9.57±0.66) μg · m-2 · h-1)>下坡位((8.26 ±1.08)μg · m-2 · h-1)>上坡位((5.59±0.74) μg · m-2 · h-1).中坡位土壤在非生长季出现多个N2O排放峰值, 根据Brumme等(1999)有关温带森林土壤N2O排放类型的划分, 可将中坡位N2O排放划分为季节事件型排放, 上、下坡位N2O排放划分为季节型排放.
2015年1月22日, 中坡位出现N2O排放最大值811.37 μg · m-2 · h-1, 结合试验地气候因素及中坡位土壤的理化特性, 可能是以下4种因素的综合作用:① WFPS:当WFPS大于60%时, 土壤内N2O产生的主要方式是反硝化作用.Zechmeister-Boltenstern等(2007)通过对51种欧洲温带森林土壤的研究发现, 大部分森林土壤N2O排放的最适WFPS均在80%以上, 2015年1月22日, 中坡位WFPS高达92.18%.而上、下坡位WFPS分别为19.9%和37.5%, 相较而言, 中坡位的WFPS有利于反硝化细菌的大量繁殖, 且更接近于Zechmeister-Boltenstern等(2007)提出的N2O排放最适WFPS值.②地温:2015年1月22日, 中坡位正午地温1.55 ℃, 夜间地温处于0 ℃以下, 昼夜气温交替使得地温在0 ℃左右反复变动, 土壤水随之在结冰-消融的状态间交替转换, 符合冻融循环发生的基本条件(王连峰等, 2007), 上、下坡位虽在地温变化上可能与中坡位相似, 但由于WFPS过低, 因而不会发生冻融循环.③营养物质:中坡位硝态氮含量为18.06 μg · g-1, 分别是下坡位的2倍(9.23 μg · g-1)及上坡位的近3倍(6.43 μg · g-1), 因而中坡位较高的硝态氮含量保证了土壤内反硝化作用的顺利进行(闵航等, 2002).Ollinger等(2002)指出当土壤C : N低于25 : 1时, 氮循环速率会进一步加快, 促进N2O排放.中坡位土壤C : N为15.59 : 1, 远低于25 : 1的临界值, 且在3个坡位间也最小, 因此, 也能促进N2O排放.④土壤质地:各坡位N2O年排放量大小与土壤粒径排序相反, 即中坡位(粉砂壤土)>下坡位(壤土)>上坡位(砂壤土).进入越冬期后土壤冻结, 土粒表面的水分会因冻结而形成隔热冰层, 冰层内未冻结水所营造的厌氧条件有利于反硝化细菌释放N2O(Teepe et al., 2001), 同时外部冰层会阻止产生的N2O向外扩散.Goossens等(2001)发现, 土质越细未冻结水含量越高, 土壤水分含量过高所形成的厌氧环境会进一步增强反硝化细菌活性, N2O产生量随之增高, 中坡位土壤粒径最小、WFPS最高, 这些因素导致了冰冻土壤内N2O的大量产生并且滞留其中, 冰层溶解后冰冻期累积的N2O集中释放, 从而出现N2O排放峰值.Brumme等(1999)的研究也发现, 在冻融交替期, 桤木森林土壤N2O的排放峰值接近500 μg · m-2 · h-1 (以N计).Luo等(2012)在德国Hglwald温带森林的研究发现, 在冻融交替期, N2O排放峰值接近800 μg · m-2 · h-1(以N计), 且峰值出现前一个月, 试验地土壤处于持续霜冻状态.Luo等(2012)认为土壤霜冻导致土壤内微生物与植物根系大量死亡, 从而为反硝化细菌提供了足够的碳、氮来源, 同时越冬期内试验地WFPS处于70%~80%之间, 为反硝化细菌提供了理想的生存环境, 加之冻融交替的作用, N2O排放峰值由此出现.
2015年9月25日, 上、下坡位土壤同时出现N2O年排放最大值.相反, 中坡位出现N2O最大吸收值(130.72 μg · m-2 · h-1), 这可能是由于采样前3天(2015年9月22—24日)采样地点累计降雨13.3 mm, 上、下坡位表层土壤的WFPS值分别为34.87%和42.12%, 可以推断降雨事件导致上、下坡位表层土壤发生干湿交替, 从而出现N2O排放峰值.中坡位植被覆盖度高, 且正处于秋季落叶期, 大量凋落物覆盖在中坡位土壤, 导致该处土壤水分蒸发量小, 加之中坡位土壤排水性较差, 因而中坡位WFPS为69.95%, 此时中坡位土壤主要发生反硝化作用.另外, 凋落物的分解提供了大量的活性有机碳, 过厚的凋落物还可能在雨水的浸润作用下进一步阻隔氧气向土壤中扩散, 从而在土壤内形成的极端厌氧环境, 这些都推动了反硝化作用彻底进行, 有利于反硝化细菌在N2O还原酶的作用下将土壤内生成的N2O还原为N2, 从而出现N2O吸收的现象.
4.3 CH4通量总体上, 油松林土壤是CH4的吸收汇, 各坡位CH4平均吸收量排序为:下坡位((77.43±14.27) μg · m-2 · h-1)>上坡位((56.16±6.76) μg · m-2 · h-1)>中坡位((43.86±9.28) μg · m-2 · h-1), 这与Smith(2000)在北欧温带针叶林所测的CH4平均吸收量相符(1.14~79.05 μg · m-2 · h-1), Castro(1992)在美国温带森林也得出类似结果.
生长季内, 上、下坡位甲烷吸收峰值的出现, 总会伴随着降雨事件的发生, Werner和Egbert(2009)发现土壤CH4吸收速率与土壤含水量呈抛物线关系, 抛物线的中点即是临界值, 低于该值, WFPS与CH4吸收速率正相关, 反之则负相关.可以推测, 试验地上、下坡位的土壤含水量均处于临界值以下, 降雨事件后土壤CH4的吸收速率随着WFPS的提升而提升, 土壤CH4的吸收峰值由此出现.中坡位土壤水分含量高于临界值, WFPS与CH4吸收速率负相关, 故降雨后中坡位CH4吸收量为坡位间最低水平, 甚至出现CH4排放的现象.大量研究发现, 温带针叶林土壤均会出现短暂的CH4排放现象(Peichl et al., 2010), 例如, 在Savage(1997)的研究中, 北美短叶松林CH4排放量高达2500 μg · m-2 · h-1.2014年11月20日, 试验地排放CH4 80.45 μg · m-2 · h-1, 在CH4排放前, 中坡位WFPS(0~20 cm)高达95%, 水分过高形成的厌氧环境有助于产生甲烷生成菌(Castro et al., 1992), 甲烷生成菌产生的甲烷会因为土壤透气性差滞留在土壤内, 等待被甲烷氧化菌再次氧化(Cai et al., 1999), 但中坡位过高的土壤含水量不利于构造甲烷氧化菌所需的好气环境(Boeckx et al., 1997), 加之冬季低温抑制了甲烷氧化菌的活性(成臣等, 2015), 因而中坡位土壤的CH4氧化吸收进程较为缓慢.由此看来, 中坡位土壤的厌氧环境一方面有助于甲烷生成菌产生CH4, 另一方面抑制了甲烷氧化菌吸收CH4的能力, 二者综合作用导致CH4在土壤内大量累积, 降水结束后随着WFPS的下降, 土壤透气性大幅增强, 土层内滞留的CH4集中排出, 暂时性的CH4排放由此形成.
5 结论(Conclusions)1) 总体来说, 温带油松林土壤是CO2、N2O的排放源, 是CH4的吸收汇.土壤温度是CO2通量季节变化的主控因素;冻融循环是导致N2O在非生长季大量排放的主要原因;上、下坡位(WFPS 30%~60%)CH4吸收峰值的出现通常伴随着降雨事件的发生, 而中坡位(WFPS 60%~95%)则在一次降雨后发生了短暂的CH4排放现象, 这表明当WFPS低于一定值时, 降雨促进土壤吸收CH4, 反之, 则抑制.
2) 坡位间土壤质地和水分的差别是引起CO2与N2O通量形成差异的主要原因.不同坡位土壤温室气体的全球增温潜势(GWP)从大到小依次是上坡位、下坡位和中坡位.
3) 本研究中, 中坡位土壤N2O通量显著高于上坡位(p < 0.01), 但未发现土壤CO2、CH4通量在坡位尺度上存在显著性差异.
Boeckx P, Ovan C, Villaralvo I. 1997. Methane oxidation in soils with different textures and land use[J]. Nutrient Cycling in Agroecosystems, 49(1/3): 91–95.
DOI:10.1023/A:1009706324386
|
Brumme R, Borken W, Finke S. 1999. Hierarchical control on nitrous oxide emission in forest ecosystems[J]. Global Biogeochemical Cycles, 13(4): 1137–1148.
DOI:10.1029/1999GB900017
|
Cai Z, Yan X. 1999. Kinetic model for methane oxidation by paddy soil as affected by temperature, moisture and N addition[J]. Soil Biology & Biochemistry, 31(5): 715–725.
|
Castro M S, Steudler P A, Melillo J M, et al. 1992. Exchange of N2O and CH4 between the atmosphere and soil in spruce-fir forests in the northeastern United States[J]. Biogeochemistry, 18(3): 119–135.
DOI:10.1007/BF00003273
|
成臣, 曾勇军, 杨秀霞, 等. 2015. 不同耕作方式对稻田净增温潜势和温室气体强度的影响[J]. 环境科学学报, 2015, 35(6): 1887–1895.
|
Corre M D, Schnabel R R, Stout W L. 2002. Spatial and seasonal variation of gross nitrogen transformations and microbial biomass in a Northeastern US grassland[J]. Soil Biology & Biochemistry, 34(4): 445–457.
|
Davidson E A, Belk E, Boone R D. 2002. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest[J]. Global Change Biology, 4(2): 217–227.
|
Davidson E A, Janssens I A, Luo Y. 2006. On the variability of respiration in terrestrial ecosystems:moving beyond Q10[J]. Global Change Biology, 12(2): 154–164.
DOI:10.1111/gcb.2006.12.issue-2
|
Fang Y T, Gundersen P, Zhang W, et al. 2009. Soil-atmosphere exchange of N2O, CO2, and CH4, along a slope of an evergreen broad-leaved forest in southern China[J]. Plant & Soil, 319(2): 37–48.
|
Goossens A, Ade V, Boeckx P, et al. 2001. Two-year field study on the emission of N2O from coarse and middle-textured Belgian soils with different land use[J]. Nutrient Cycling in Agroecosystems, 60(1/3): 23–34.
DOI:10.1023/A:1012695731469
|
Hirobea M, Iwatsubo G. 1998. Spatial variability of soil nitrogen transformation patterns along a forest slope in a Cryptomeria japonica D.Don plantation[J]. European Journal of Soil Biology, 34(3): 123–131.
DOI:10.1016/S1164-5563(00)88649-5
|
Hutchinson G L. 2004. Improved soil cover method for field measurement of nitrous-oxide fluxes[J]. Soil Science Society of America Journal, 45: 311–316.
|
Lal R. 2004. Soil carbon sequestration to mitigate climate change[J]. Geoderma, 123(1-2): 1–22.
DOI:10.1016/j.geoderma.2004.01.032
|
李玉田, 庞军柱, 彭长辉, 等. 2017. 秦岭火地塘林区不同海拔森林土壤NO通量[J]. 东北林业大学学报, 2017, 45(2): 42–49.
|
Luo G J, Brüggemann N, Wolf B, et al. 2012. Decadal variability of soil CO2, NO, N2O, and CH4 fluxes at the Höglwald Forest, Germany[J]. Biogeosciences, 9(5): 1741–1763.
DOI:10.5194/bg-9-1741-2012
|
Mcclain M E, Boyer E W, Dent C L, et al. 2003. Biogeochemical hot spots and hot moments at the interface of terrestrial and aquatic ecosystems[J]. Ecosystems, 6: 301–312.
DOI:10.1007/s10021-003-0161-9
|
Mcswiney C P, Mcdowell W H, Keller M. 2001. Distribution of nitrous oxide and regulators of its production across a tropical rainforest catena in the Luquillo Experimental Forest, Puerto Rico[J]. Biogeochemistry, 56(3): 265–286.
DOI:10.1023/A:1013191010244
|
Merino A, Perezbatallon P, Macias F. 2004. Responses of soil organic matter and greenhouse gas fluxes to soil management and land use changes in a humid temperate region of southern Europe[J]. Soil Biology & Biochemistry, 36(6): 917–925.
|
闵航, 陈中云, 吴伟祥, 等. 2002. 碳、氮物质对水稻田土壤甲烷氧化活性影响的研究[J]. 环境科学学报, 2002, 22(1): 70–75.
|
Ollinger S V, Smith M L, Martin M E, et al. 2012. Regional variation in foliar chemistry and N cycling among forests of diverse history and composition[J]. Ecology, 83(2): 339–355.
|
Osaka K, Ohte N, Koba K, et al. 2015. Hydrologic controls on nitrous oxide production and consumption in a forested headwater catchment in central Japan, Journal of Geophysical Research Biogeosciences[J]. 111(G1): 277-282
|
庞军柱, 王效科, 牟玉静, 等. 2010. 黄土高原苹果园土壤N2O排放研究[J]. 环境科学学报, 2010, 30(12): 2518–2525.
|
Peichl M, Arain M A, Ullah S, et al. 2010. Carbon dioxide, methane, and nitrous oxide exchanges in an age sequence of temperate pine forests[J]. Global Change Biology, 16(8): 2198–2212.
|
Rosenkranz P, Seufert G, Butterbach B K, et al. 2006. N2O, NO and CH4 exchange, and microbial N turnover over a Mediterranean pine forest soil[J]. Biogeosciences, 3(2): 21–133.
|
Russell C A, Voroney R P, Black T A, et al. 1998. Carbon dioxide efflux from the floor of a boreal aspen forest.Ⅱ.Evaluation of methods--verification by infra-red analysis of a dynamic closed chamber[J]. Canadian Journal of Soil Science, 78(2): 301–310.
DOI:10.4141/S97-054
|
Ryota K, Seiichi O, Shigehiro I, et al. 2008. Spatial structures of N2O, CO2, and CH4 fluxes from Acacia mangium plantation soils during a relatively dry season in Indonesia[J]. Soil Biology & Biochemistry, 40(12): 3021–3030.
|
Savage K, Moore T R, Crill P M. 1997. Methane and carbon dioxide exchanges between the atmosphere and northern boreal forest soils[J]. Journal of Geophysical Research Atmospheres, 102(24): 29279–29288.
|
Schmidt J, Seiler W, Conrad R. 1987. Emission of nitrous oxide from temperate forest soils into the atmosphere[J]. Journal of Atmospheric Chemistry, 6(1): 95–115.
|
Smith K A, Dobbie K E, Ball B C, et al. 2000. Oxidation of atmospheric methane in Northern European soils, comparison with other ecosystems, and uncertainties in the global terrestrial sink[J]. Global Change Biology, 6(7): 791–803.
DOI:10.1046/j.1365-2486.2000.00356.x
|
Teepe R, Brumme R, Beese F. 2001. Nitrous oxide emissions from soil during freezing and thawing periods[J]. Soil Biology & Biochemistry, 33(9): 1269–1275.
|
Ullah S, Moore T R. 2015. Biogeochemical controls on methane, nitrous oxide, and carbon dioxide fluxes from deciduous forest soils in eastern Canada, Journal of Geophysical Research: Biogeosciences[J]. 116(G3): 3010
|
Wang C, Bond L B, Gower S T. 2003. Soil surface CO2 flux in a boreal black spruce fire chronosequence[J]. Journal of Geophysical Research, 108(D3): 5–8.
|
Werner B, Egbert M. 2009. Reappraisal of drying and wetting effects on C and N mineralization and fluxes in soils[J]. Global Change Biology, 15(4): 808–824.
DOI:10.1111/gcb.2009.15.issue-4
|
West T O, Marland G. 2002. A synthesis of carbon sequestration, carbon emissions, and net carbon flux in agriculture:comparing tillage practices in the United States[J]. Agriculture Ecosystems & Environment, 91(1): 217–232.
|
王连峰, 蔡延江, 解宏图. 2007. 冻融作用下土壤物理和微生物性状变化与氧化亚氮排放的关系[J]. 应用生态学报, 2007, 18(10): 2361–2366.
|
Xu W, Jiang Y L, Jia B R, et al. 2010. Comparison of soil respiration among three temperate forests in Changbai Mountains, China[J]. Canadian Journal of Forest Research, 40(4): 788–795.
DOI:10.1139/X10-010
|
Zechmeister-Boltenstern S, Schaufler G, Kitzler B. 2007. NO, NO2, N2O, CO2 and CH4 fluxes from soils under different land use:temperature sensitivity and effects of soil moisture[J]. Geophysical Research Abstracts, 8: 7968.
|