 2019, Vol. 39
2019, Vol. 39



原位生物修复被认为是减少地下水体污染的一项重要技术(Verardo et al., 2017), 其中, 碳源的选择至关重要(Li et al., 2015; 赵阅坤, 2016).乳化油作为一种新兴缓释碳源, 具有成本低廉、易形成反应带、缓慢释放电子等优点(丁琳洁, 2016), 因而受到越来越多国内外学者的关注.目前, 国内外针对乳化油在地下水污染原位修复中的研究与应用, 已涉及到硝酸盐污染(Hunter, 2001; Vujasinovic et al., 2014)、氯代溶剂污染(Newman et al., 2006; Harkness et al., 2013; Liang et al., 2013; Hiortdahl et al., 2014; Tsai et al., 2014)、高氯酸盐污染(Borden, 2007; Wang et al., 2013)、六价铬等重金属污染(Lindow et al., 2005; Zhang et al., 2015; 生贺等, 2015; Wen et al., 2017)及放射性铀污染(Tang et al., 2013; Waston et al., 2013; Deng et al., 2016)等领域.然而在实际应用过程中, 出现了含水层介质的渗透性损失甚至堵塞现象, 导致修复效率的降低或失效.对此, 何宝南等(2018)在乳化油基础上制备了粒径更小的纳米乳化油, 从物理截留角度探讨了其对含水层介质渗透性的影响.然而, 对于纳米乳化油降解污染物过程中产气及微生物增殖代谢等引起的堵塞还关注较少.
已有相关研究报道, 微生物降解污染物过程中的产气及微生物增殖代谢对含水层所造成的堵塞现象不可忽视.例如, Hunter(2001)在以植物油为碳源去除地下水硝酸盐污染时发现多孔介质渗透性损失约27%, 推测认为可能是微生物脱氮产生N2和CO2引起;Borden(2007)在利用乳化油增强地下水中高氯酸盐和氯化溶剂的原位生物降解的现场中试试验中指出注入井的渗透性下降了39%~91%(平均68%), 可能是由于气体产生和生物量增长造成.尽管上述有关乳化油在地下水原位修复的研究中均提到了产气和微生物增殖可能会导致渗透性的下降, 但依然停留在推测阶段, 且仅显示产气和微生物增殖代谢造成渗透性损失的最终状态, 忽略了乳化油发酵及污染物降解过程中产气及微生物生长代谢及产物阶段性产量的增减特征和累积过程, 以及不同堵塞作用共存下对于堵塞的协同贡献和主导因素.
为此, 本文选取降解过程中既有产气又有微生物代谢的典型污染物硝酸盐为代表, 以纳米乳化油(Nano Emulsified Vegetable Oil, Nano-EVO)为碳源, 采用市售反硝化菌剂接种微生物开展硝酸盐降解的模拟实验研究, 通过监测硝酸盐降解过程中产气和微生物的增殖代谢过程, 模拟评估纳米乳化油修复地下水硝酸盐过程中产气及微生物增殖代谢对多孔介质的堵塞作用, 为探索行之有效的缓解措施提供依据.
2 材料与方法(Materials and methods) 2.1 实验材料纳米乳化油配制原料:大豆油(金龙鱼, 市售)、食品级乳化剂(吐温80和斯盘80, 天津光复精细化工研究院).硝酸盐氮(NO3--N)、亚硝酸盐氮(NO2--N)和氨氮(NH4+-N)检测试剂:氨基磺酸、盐酸、对-氨基苯磺酰胺、N-(1-萘基)-乙二胺二盐酸盐、酒石酸钾钠和纳氏试剂(分析级).碳酸根和碳酸氢根检测药品:无水碳酸钠和盐酸(0.025 mol·L-1), 购自国药化学试剂有限公司.市售反硝化菌剂主要是由假单胞菌属、产检杆菌属、科奈瑟菌科、红螺菌科、芽孢杆菌科和纤维粘菌科等组成的反硝化菌群, 购自上海甘度环境工程有限公司.微生物计数:3M PetrifilmTM菌落总数测试片(美国3M公司).胞外聚合物提取及蛋白质、多糖测定试剂:氯化钠、考马斯亮蓝G250染色液、浓硫酸和苯酚.
2.2 实验方法 2.2.1 纳米乳化油制备由课题组何宝南等(2018)的方法制备而成:借助升温相转化技术, 将大豆油、吐温80、斯盘80和去离子水按照一定比例混合, 经电动搅拌、超声及恒温水浴加热等步骤处理后制得乳白色的纳米乳化油, 平均粒径大小为255.1 nm, 稳定性动力学指数为0.5, COD释放为4.72×105 mg·L-1.
2.2.2 静态批实验设计设计开展纳米乳化油降解硝酸盐的静态批实验, 实验装置参照课题组连玉倩等(2018)的设计装置(图 1), 其中, 反应器用于反硝化菌剂降解硝酸盐氮, 主要监测瓶中硝酸盐氮的降解及微生物的生长代谢情况, CO2吸收瓶用于吸收碳源厌氧发酵过程中产生的二氧化碳, 以测定和计算出系统的CO2产气量, U型管用于测定硝氮降解过程中N2及其他类型非溶性气体的产气情况, 通过记录U型管的液压差来获取该部分气体的总产气量.反应溶液体积控制在1200 mL, 采用一次性加入3 mL纳米乳化油、0.75 g反硝化菌剂、硝酸盐氮溶液周期性加入的方式启动, 其中, 硝酸盐氮的初始质量浓度设置为50 mg·L-1, 当其降至不变时记作一个周期.实验在常温、避光的条件下进行, 包含3个平行组.
 |
| 图 1 实验装置示意图 Fig. 1 Schematic diagram of experimental set-up |
硝酸盐氮(NO3--N)、亚硝酸盐氮(NO2--N)和氨氮(NH4+-N)分别采用紫外分光光度法(B)、N-(1-萘基)-乙二胺光度法(A)和纳氏试剂光度法(A)完成测定, 参照《水和废水的监测和分析方法(第四版)》(魏复盛等, 2002).
CO2产气量通过电位滴定法测定碱度来量化, 根据溶液pH值达到8.3和4.4两个滴定终点时所消耗的标准盐酸溶液用量, 计算出CO2吸收瓶中碳酸根和碳酸氢根含量;CH4、N2O等的浓度采用气相色谱仪进行检测.
当V1<1/2V2时:
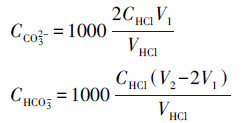
|
(1) |
当V1=0时:

|
(2) |
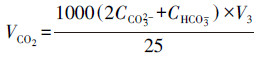
|
(3) |
式中, CCO32-为瓶中碳酸根浓度(mol·L-1);CHCO3-为瓶中重碳酸根浓度(mol·L-1);CHCl为实验所用盐酸标准溶液浓度(mol·L-1);VHCl为盐酸标准溶液消耗量(mL);V1为滴定至pH=8.3时盐酸标准溶液消耗量(mL);V2为滴定至pH=4.4时盐酸标准溶液消耗量(mL);V3为测样体积, 取值为25 mL.
微生物数量基于连续稀释和3M PetrifilmTM菌落测试片计数建立的OD600-微生物浓度对应关系来表征.
胞外聚合物(Extracellular Polymeric Substances, EPS)的提取及其组分的测定:EPS采用热提法进行提取(吕英, 2007), 对所取样品进行预处理, 在7300 r·min-1(6000 g)下离心10 min(4 ℃)后弃去上清液, 加入0.9% NaCl缓冲液恢复体积.将预处理后的样品在80 ℃下水浴加热30 min, 再在8000 r·min-1下离心10 min, 取上清液过0.22 μm微孔滤膜即为EPS提取液.EPS的主要成分包括蛋白质(Protein, PN)和多糖(Polysaccharide, PS)(Flemming et al., 2010), 约占总量的75%~80%, 故本实验以蛋白质和多糖总量来表征EPS含量, 多糖及蛋白质分别依据浓硫酸-苯酚法(中华人民共和国农业部, 2008)和考马斯亮蓝法(Bradford, 1976)的原理采用分光光度法在490 nm、595 nm下完成测试(HACH, DR6000).
2.2.4 多孔介质一维柱概念模型与地下水系统假设为了进一步探讨实际修复过程中出现的含水层堵塞现象, 前期弱化维度的设置, 从简单的一维条件开始探讨.基于前期减小乳化油粒径以缓解多孔介质渗透性损失的实验研究成果(何宝南等, 2018), 建立纳米乳化油在多孔介质中的一维柱概念模型, 其具体参数如表 1所示.假设该模型分别采用中砂和细砂两种介质装填, 介质在沿柱的水平横截面都是均匀各向同性分布的, 每种介质中均注入15 mL的纳米乳化油, 分别占孔隙体积的2.5%和2.8%, 柱体内部为等速的均匀流场, 且不考虑流动时边界条件带来的效应.
| 表 1 多孔介质一维柱实验概念模型参数 Table 1 The basic parameters of one-dimensional column conceptual model |
同时, 假设模型所对应的地下水系统在潜水深度以下约50 m处, 实际水流速度ν为常数, 水温20 ℃(批实验是在此温度下进行), 压力为0.5 MPa(≈ 5 atm);假设含水层系统为封闭体系, 即含水层系统与外界大气之间无气体交换, 默认此系统内产生的气体部分溶于水, 而未溶解的气体则会不断地累积致使含水层多孔介质发生堵塞.
2.2.5 产气的溶解性和渗透性变化计算方法受压力变化影响, 水中部分溶解的气体在气液平衡作用下可能会被释放, 同时, 未溶解气体的溶解度也可能随压力而变化.在某种程度上, 这部分剩余气体将和水一起占据孔隙体积, 使多孔介质的初始孔隙率(ninit)降低并导致堵塞.以下通过亨利定律和理想气体状态方程来计算平衡系统中气体的溶解度:
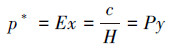
|
(4) |
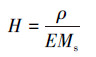
|
(5) |
式中, p*为气相中溶质的平衡分压(kPa);E为亨利常数(kPa), 与温度相关;x为溶质在液相中的摩尔分率;c为气体在水中的溶解度(kmol·m-3);H为溶解系数(kmol·m-3·Pa-1);P为实际气压(kPa);y为气相中的溶质百分比;ρ为水的密度(g·cm-3);Ms为水的摩尔质量, 取值为18 g·mol-1.
理想气体状态方程pV=nRT, 用密度表示该关系即为:
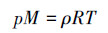
|
(6) |
式中, p为理想气体压强(Pa);M为理想气体的摩尔质量(g·mol-1);ρ为气体密度(kg·m-3);R为理想气体常数, 取值为8.314 Pa·m3·mol-1·K-1;T为体系的热力学温度(K).
采用Kozeny-Carman(K-C)方程(Henderson et al., 2011; Jeen et al., 2012)来量化产气对多孔介质渗透性的影响:
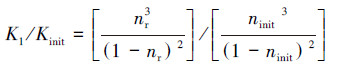
|
(7) |
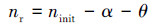
|
(8) |
式中, Kinit为初始渗透系数(m·d-1);K1为产气影响后的渗透系数(m·d-1);ninit为多孔介质的初始孔隙率;nr为产气后的残余孔隙率;α为气体含量, 等于气体体积/总体积;θ为水含量, 等于水体积/总体积.
2.2.6 微生物细胞生物量和胞外聚合物引起的渗透性变化预测模型Clement模型(Clement et al., 1996):该模型没有将生物膜细分为细菌和胞外聚合物, 对多孔介质中微生物在孔隙中的任何特定分布也没有作出具体假设, 仅使用固体颗粒表面微生物量的数据预测微生物致使的渗透性下降.具体公式如下:
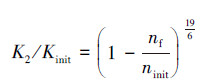
|
(9) |
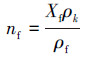
|
(10) |
式中, Kinit为初始渗透系数(m·d-1);K2为微生物影响后的渗透系数(m·d-1);ninit为初始孔隙率;nf为固体颗粒表面的微生物所占孔隙的体积分数;Xf为每单位质量含水层颗粒上微生物细胞的干重, 依据与OD600值的关系(y=0.951x+0.0722, R2=0.915)(唐远江等, 2016)得到;ρk为含水层的颗粒密度(g·cm-3);ρf为微生物的密度.假设微生物细胞皆为球形, 其直径为10-4 cm, 细胞重为10-13 g, 其中, 蛋白含量为55%(杨靖等, 2011).
Pham模型(Pham et al., 2018):该模型从孔隙网络尺度出发, 将多孔介质视为最简单的平行毛细管束, 对于均匀涂层形式, 当生物量积累时, 生物膜均匀地覆盖每个毛细管的内壁, 导致孔隙半径从r0减小到r.具体公式如下:
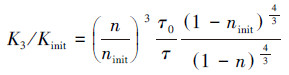
|
(11) |
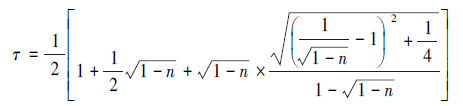
|
(12) |
式中, Kinit为初始渗透系数(m·d-1);K3为管孔半径减小后的渗透系数(m·d-1);τ表示毛细管的弯曲程度;ninit为多孔介质的初始孔隙度;n为多孔介质的几何尺寸受到生物量积累的影响后的孔隙度;假设生物膜质量(SV)由EPS质量(SEPS)和细菌质量(SCell)组成(蒲晓芬等, 2005):SV=SEPS+SCell, 借由蛋白含量将细菌和胞外聚合物两者定量处理为归一化表面负荷.
Kozeny Grain(KG)模型(Abbasi et al., 2018):该模型考虑颗粒介质中孔隙空间的不规则性, 设定两种生物膜的生长形式, 将孔隙半径减小导致的渗透性降低和孔隙堵塞导致的渗透性降低看作多孔介质整体渗透性降低的可能机制.
① 生物膜覆盖砂粒表面及与砂粒接触的颗粒包覆模型:
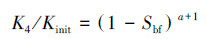
|
(13) |
式中, Kinit为初始渗透系数(m·d-1);K4为孔隙半径减小后的渗透系数(m·d-1);Sbf为生物膜饱和度;a为饱和指数, 当0<Sbf<0.8时取a=1.5(Abbasi et al., 2018).
② 生物膜从颗粒表面开始向孔隙空间垂直生长的孔隙填充模型(Kleinberg et al., 2003):
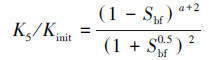
|
(14) |
式中, Kinit为初始渗透系数(m·d-1);K5为孔隙堵塞后的渗透系数(m·d-1).
3 结果与讨论(Results and discussion) 3.1 “三氮”降解情况图 2显示在实验Ⅰ~Ⅳ的4个周期内硝酸盐氮浓度均有所降低, 表明在纳米乳化油刺激下, 反硝化菌对硝氮具有较强的降解能力, 4个周期的硝氮降解率分别为92.87%、91.79%、97.38%和78.01%, 共降解硝氮233.23 mg, 总去除率为90.23%.其中, 前3个周期显示出了纳米乳化油碳源的降解优势, 反硝化菌的硝氮降解率均在90%以上, 随着碳源不断消耗, 第Ⅳ周期的降解率有所减少.
 |
| 图 2 纳米乳化油刺激下的“三氮”变化 Fig. 2 Concentration variation of NO3--N, NO2--N and NH4+-N under the stimulation of Nano-EVO |
在前两个周期, 系统中都有不同程度的亚硝氮积累, 且在NO3--N被完全降解时累积量达到最大, 随后逐渐下降至检出限附近并趋于稳定, 无积累.该结果与董晓莹(2017)对反硝化过程的研究结论相一致, 当系统中同时存在NO3--N和NO2--N两种电子受体时, 由于硝酸盐对于碳源的竞争力大于亚硝酸盐对于碳源的竞争力, 反硝化菌会优先还原NO3--N, 导致NO2--N的暂时积累.
系统中氨氮浓度在Ⅰ周期开始发生小幅升高并积累, Ⅱ、Ⅲ周期的氨氮含量水平在7.35~18.35 mg·L-1之间稳定波动, 第Ⅳ周期略呈现下降趋势.
3.2 产气及代谢产物动态变化纳米乳化油降解硝酸盐过程中产生气体的主要来源包括微生物的新陈代谢(无氧呼吸作用产生CO2)、碳源在微生物作用下的厌氧发酵(CH4、CO2等)及硝酸盐氮降解产气(反硝化作用下N2的生成).经过实验装置中的CO2吸收瓶(连玉倩等, 2018)后, 检测所得CH4和N2O浓度所占百分比<0.001%, 可忽略不计, 故反应体系中的主要产气类型为CO2和N2.如图 3a所示, 在整个实验周期中, 累积产气198.83 mL, 其中, CO2共165.12 mL, N2共33.71 mL, 降解1 mg硝氮对应的平均CO2产气量为0.71 mL, 平均N2产气量为0.14 mL, 且随着周期的进行, 纳米乳化油不断被消耗, 阶段产气量整体呈现下降趋势.
 |
| 图 3 周期内产气和微生物代谢产物的动态变化 Fig. 3 The dynamic change of gases production and metabolites content during the research cycles |
微生物的生长情况如图 3b所示, 在纳米乳化油降解硝酸盐静态批试验的4个周期中, 受接种多种细菌后可能会发生不同菌种之间的竞争性抑制生长的影响, 反硝化菌群的生长曲线整体呈下降趋势, 一些不适应环境的菌逐渐死亡, 部分菌则存活下来并成为优势菌群.
微生物代谢产物主要来源于细菌的分泌、细菌表面物质的脱落、细菌死亡后的自溶及对周围环境物质的吸附所构成的EPS.与产气相反, EPS的阶段产量则呈现上升趋势, 主要表现在Ⅱ~Ⅳ周期, 蛋白质和多糖相较于Ⅰ周期均显示出明显增长, 微生物倾向于产生更多的胞外聚合物来保护自己免受外界环境的不利影响(Yang et al., 2013; More et al., 2014; 刘瑶, 2017).在4个周期内, 系统共计产生胞外聚合物186.65 mg, 其中, 蛋白质149.37 mg, 阶段性的产量占比约为67.09%~93.86%, 多糖37.28 mg, 阶段性的产量占比约为6.14%~32.91%, 降解1 mg硝氮对应的平均蛋白产量为0.64 mg, 平均多糖产量为0.16 mg, PN/PS比为4.01.有研究表明, 基质种类对EPS产量的影响很大(Sponza, 2003), 这说明系统中的外加碳源纳米乳化油对EPS组分含量的不同可能具有一定作用.在整个实验阶段, 测得的PN含量要远大于PS, 这表明蛋白在抵御50 mg·L-1硝氮浓度的环境中起到了更关键的作用, 菌群分泌了更多的胞外蛋白来保护自身.且PN中可能包含了许多NO3--N的还原酶, 能够将NO3--N(+5)还原为N2(0), 重复性的周期开展及硝氮浓度的人为提升, 引导刺激产生了更多的还原酶, 同时也产生了更多的EPS.
3.3 多孔介质堵塞评估 3.3.1 产气对多孔介质渗透性的影响在整个实验中, 未溶解气体被视为“废气”, 如果这些“废气”没有及时有效地排出, 多孔介质的渗透性会受到影响甚至发生严重下降.本研究利用2.2.4节中的假设条件、20 ℃下CO2和N2对应的亨利常数及批实验得到的产气量, 基于产气的溶解性, 根据式(4)~(6)计算气体溶解度, 得到CO2和N2在地下水系统中的溶解度分别为0.941 L·L-1和0.017 L·L-1;再利用批实验中3 mL纳米乳化油原位处理硝酸盐过程中的产气量进行等比换算, 得到一维模拟柱中注入15 mL纳米乳化油时的产气情况, 具体如表 2所示.
| 表 2 一维模拟柱理论计算的产气情况 Table 2 The theoretical gas production of one-dimensional simulated column |
对一维模拟柱内的未溶解气体, 通过K-C方程进行渗透性下降情况的预估, 结果如图 4所示.经过2个周期左右的时间, 系统中的累积气体产量最终导致了中/细砂内的渗透性完全损失.与此同时, 可以发现细砂介质中的渗透性比中砂中降低得更快, 这与多孔介质的结构密切相关(杜磊, 2016).
 |
| 图 4 气体对一维模拟柱内多孔介质渗透性的影响 Fig. 4 Effect of gases production on permeability of porous media in one-dimensional simulated column |
对于因孔隙中胞外聚合物存在而堵塞的砂柱, 若仅对细菌进行量化, 会大大低估多孔介质中的生物量(Thullner, 2010).本研究以实验所得细胞生物量和胞外聚合物为基础, 分别采用Clement、Pham及KG模型评估由生物量和胞外聚合物造成的一维模拟柱的渗透性损失.如图 5所示, 3种模型下的曲线变化趋势相同, 均呈“对勾型”曲线, 即在一维模拟砂柱中, 多孔介质的渗透性均在系统开始运行阶段就出现骤降, 而后有所回升, 恢复后的K要小于砂柱的初始渗透系数.在实验前期(0~5 d), EPS浓度较低(图 3), 多孔介质渗透性的骤降主要是由初始阶段较高数量的反硝化菌引起的, 此时微生物可能位于砂粒接触处或孔颈处, 即使仅占孔隙的一小部分, 也会造成显著的堵塞(Xia et al., 2016).实验中期(5~30 d), EPS的含量较前期提高, 但一维模拟柱的渗透性却出现小幅回升, 表明该时期微生物数量的下降对渗透性起到主要影响.实验后期(30~58 d), 与微生物变化相反, EPS产量呈明显增长趋势, 但砂柱的渗透性仍在逐渐回升, 说明EPS产量尽管在一定程度上减缓了渗透性的恢复, 但其增长对渗透性没有显著影响, 而微生物细胞数量的增减在大部分实验期间对渗透性损失起了主导作用.
 |
| 图 5 3种模型对微生物引起的渗透性变化预测结果对比 Fig. 5 Comparison of three models (Clement/Pham/KG) for predicting microbial-induced permeability changes |
上述3种模型的评估结果显示(图 5)其预测的渗透性损失由大到小依次为:Pham模型>KG模型>Clement模型, 在中砂介质中, 造成的最终渗透性损失分别为31.10%、29.90%和10.87%, 在细砂介质中, 造成的最终渗透性损失分别为48.32%、30.80%和12.77%, 细砂的渗透性损失要高于中砂.这与Thullner等(2004)的研究结果一致, 对于颗粒直径0.09 ~1.0 mm的多孔介质而言, 细颗粒材料比粗颗粒材料更容易堵塞.
为了验证本研究预测结果的可靠性, 选取部分案例的计算值进行对照.如表 3所示, Clement模型、Pham模型及KG模型在不同案例(杨靖, 2011;Pham et al., 2018; Abbasi et al., 2018)的应用中均显示出了较好的模拟效果, 预测值与实测值接近.其中, Abbasi等(2018)在其研究中将一维柱实验的数据用于二维流体动力学模型的堵塞模拟, 这说明低维度中实际蕴含着大量可利用的有关高维度的信息, 若已知相关流体动力参数(如密度、流速、动力粘度等), 可以通过在低维度条件下获取的相关数据来模拟高维度下的流动和溶质运移.这种方法也已在生物堵塞的相关研究中得到应用, 如Deng等(2013)从距离柱状反应器表面11 μm深处拍摄共聚焦显微镜图像获得生物膜结构用于二维流体动力学模拟, 其三维孔隙度则是通过叠加二维共聚焦图像的孔隙度计算得出.
| 表 3 微生物堵塞模型预测情况 Table 3 Prediction results of microbial clogging models |
将不同案例与本研究预测的K/K0值两两对比发现, 本研究的预测值整体要比案例中的数值低一些, 其中, KG模型的预测结果是最为接近的.考虑到实验条件如维度、介质等存在一定差异, 本文中3种模型所预测的渗透性变化结果对后续研究纳米乳化油降解硝酸盐过程中的堵塞仍具有一定的参考价值和借鉴意义.
3.3.3 产气及微生物增殖代谢对堵塞的贡献本研究以相对贡献率的占比来体现系统中造成堵塞的主控因素, 基于多孔介质一维模拟柱得到的因产气和微生物增殖代谢造成的渗透性损失结果, 在仅考虑这两种堵塞作用存在的情况下, 计算得出产气和微生物增殖代谢中砂介质渗透性损失的贡献率分别为76.28%~90.20%和9.80%~23.72%, 对细砂介质的渗透性损失贡献率分别为67.42%~88.68%和11.32%~32.58%.
由此来看, 在含水层与外界无气体交换的封闭系统中, 纳米乳化油原位修复硝酸盐过程中气体的产生和累积对含水介质渗透性损失的贡献相较于微生物增殖代谢更大, 表明地下水含水层系统的实际埋藏条件至关重要, 其决定了含水层内累积的气体是否与外界大气发生交换或释放, 从而影响了产气对多孔介质造成的堵塞程度.在PRB修复的相关研究中, 已有学者关注了这个问题, 如Birke(2003)在其研究中指出, 在开放系统中, 气体可以通过气泡聚结和浮力作用自然排出, 而在封闭系统中, 气体的高产出率和排气的受限可能会使气体积聚从而导致PRB失效;Zhang等(2005)在研究利用零价铁PRBs修复TCE污染时指出, 可以通过设置排气口来避免因气体截留而造成反应墙渗透系数的下降幅度超过一个数量级;Kamolpornwijit等(2006)的实验和建模结果显示, 气体排放对于保持封闭系统PRBs的流量和性能尤其重要;Henderson等(2011)在研究固体沉淀和产气对ZVI PRBs渗透性的影响时表明渗透性损失大部分归因于气体, 在设计PRBs时应实现系统的有效排气使气体逸出以防止渗透性的过度降低.在这些研究中, 虽然体系的开放均被涉及, 但没有人对系统的开启程度进行定义和量化.
分别取含水层系统开启程度为0(封闭体系)、25%、50%、75%、100%(开放体系)时产气和微生物增殖代谢的平均贡献率预估值作图, 以便观测实际含水层渗透性损失贡献率可能分布的范围(图 6), 结果发现, 随着含水层开启程度的增加, 渗透性损失中由产气造成的贡献占比会下降, 而由微生物造成的贡献占比则会提高, 且在中砂介质中这种转变更为明显.在文中的假设条件下, 柱子作为准一维系统运行, 水仅沿着柱子的长度流动, 忽略了孔隙和宏观尺度上多孔介质流场的多维性, 但考虑在实际含水层中, 地下水系统情况还会受其他多种因素, 如多孔介质、微生物生长代谢情景、水动力条件、环境基质等控制, 很难达到文中某些假定的理想条件, 且涉及到多维流场中的生物堵塞, 其水流运动和溶质运移过程对于边界条件更为敏感, 意味着对多维流场的生物堵塞研究比一维流场的情况更加复杂, 对产气和微生物增殖代谢引起的渗透性损失预测以及贡献率的确切数值仍应视不同含水层具体的实际情况而定.
 |
| 图 6 渗透性损失的贡献率随含水层系统开启程度的变化 (a.细砂, b.中砂) Fig. 6 Contributions of permeability loss varied with the openness degree of aquifer systems |
1) 纳米乳化油作为碳源能够有效降解硝酸盐氮污染, 4个周期反硝化菌剂的硝氮总去除率达90.23%.在整个研究周期中, 产气和代谢产物展现出相反的增减趋势, 以CO2及N2为主要产气成分的周期产气量呈下降趋势, 降解1 mg硝氮对应的平均CO2产气量为0.71 mL, 平均N2产气量为0.14 mL;以蛋白质及多糖为主要成分的微生物代谢产物胞外聚合物产量则呈明显递增趋势, 降解1 mg硝氮对应的平均蛋白产量为0.64 mg, 平均多糖产量为0.16 mg.
2) 在含水层为封闭体系, 不存在与外界气体交换的前提下, K-C方程预估产气在2个周期内导致一维模拟柱中/细砂介质内的渗透性完全损失;微生物增殖代谢对多孔介质渗透性损失影响预估结果为Pham模型>KG模型>Clement模型, 在中砂中造成的最终渗透性损失分别达31.10%、29.90%、10.87%, 在细砂中造成的最终渗透性损失分别达48.32%、30.80%、12.77%, 细砂中的渗透性损失均要高于中砂.
3) 在封闭体系, 仅考虑两种堵塞作用存在的条件下, 气体的产生和累积对纳米乳化油原位修复硝酸盐过程中含水介质渗透性损失的贡献较微生物增殖代谢要更大, 是造成堵塞的主导因素, 随着含水层开启程度的增加, 由产气造成的贡献占比会下降, 由微生物造成的贡献占比则会提高.因此, 探索增强其开放状况是缓解堵塞的有效途径之一.
Abbasi B, Ta H X, Muhunthan B, et al. 2018. Modeling of permeability reduction in bioclogged porous sediments[J]. Journal of Geotechnical and Geoenvironmental Engineering, 144(4): 04018016. DOI:10.1061/(ASCE)GT.1943-5606.0001869 |
Birke V.2003.Mainstream and lessons learned at nine German PRB sites over 5-years—an interim report[C].RTDF Meeting Proceedings. Niagara Falls, New York
|
Bradford M M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding[J]. Analytical Biochemistry, 112: 195-203. |
Clement T P, Hooker B S, Skeen R S. 1996. Macroscopic models for predicting changes in saturated porous media properties caused by microbial growth[J]. Groundwater, 34(5): 934-942. DOI:10.1111/j.1745-6584.1996.tb02088.x |
Borden R C. 2007. Concurrent bioremediation of perchlorate and 1, 1, 1-trichloroethane in an emulsified oil barrier[J]. Journal of Contaminant Hydrology, 94: 13-33. DOI:10.1016/j.jconhyd.2007.06.002 |
Deng W, Cardenas M B, Kirk M F, et al. 2013. Effect of permeable biofilm on micro- and macro-scale flow and transport in bioclogged pores[J]. Environmental Science & Technology, 47(19): 11092-11098. |
Deng Y, Zhang P, Qin Y J, et al. 2016. Network succession reveals the importance of competition in response to emulsified vegetable oil amendment for uranium bioremediation[J]. Environmental Microbiology, 18(1): 205-218. DOI:10.1111/1462-2920.12981 |
丁琳洁.2016.Y-ESO强化原位生物反应带修复硝基苯污染含水层研究[D].长春: 吉林大学 http://cdmd.cnki.com.cn/Article/CDMD-10183-1018279302.htm
|
董晓莹.2017.部分反硝化过程亚硝氮积累的控制条件及种群特性研究[D].西安: 西安建筑科技大学 http://cdmd.cnki.com.cn/Article/CDMD-10703-1017736831.htm
|
杜磊.2016.含水层回灌过程中的气相堵塞实验研究[D].济南: 济南大学 http://cdmd.cnki.com.cn/Article/CDMD-10427-1016257869.htm
|
Flemming H C, Wingender J. 2010. The biofilm matrix[J]. Nature Reviews Microbiology, 8(9): 623-633. DOI:10.1038/nrmicro2415 |
Harkness M, Fisher A. 2013. Use of emulsified vegetable oil to support bioremediation of TCE DNAPL in soil columns[J]. Journal of Contaminant Hydrology, 151: 16-33. DOI:10.1016/j.jconhyd.2013.04.002 |
何宝南, 何江涛, 王菲, 等. 2018. 纳米乳化油缓解多孔介质渗透性损失的实验研究[J]. 环境科学学报, 38(1): 267-274. |
Henderson A D, Demond A H. 2011. Impact of solids formation and gas production on the permeability of ZVI PRBs[J]. Journal of Environmental Engineering, 137(8): 689-696. DOI:10.1061/(ASCE)EE.1943-7870.0000383 |
Hiortdahl K M, Borden R C. 2014. Enhanced reductive dechlorination of tetrachloroethene dense nonaqueous phase liquid with EVO and Mg(OH)2[J]. Environmental Science & Technology, 48(1): 624-631. |
Hunter W J. 2001. Use of vegetable oil in a pilot-scale denitrifying barrier[J]. Journal of Contaminant Hydrology, 53(1): 119-131. |
Jeen S W, Amos R T, Blowes D.W. 2012. Modeling gas formation and mineral precipitation in a granular iron column[J]. Environmental Science & Technology, 46(12): 6742-6749. |
Kamolpornwijit W, Liang L. 2006. Investigation of gas production and entrapment in granular iron medium[J]. Journal of Contaminant Hydrology, 82(3): 338-356. |
Kleinberg R L, Flaum C, Griffin D D, et al. 2003. Deep sea NMR:Methane hydrate growth habit in porous media and its relationship to hydraulic permeability, deposit accumulation, and submarine slope stability[J]. Journal of Geophysical Research:Solid Earth, 108(B10): 2508. |
Li W W, Yu H Q. 2015. Electro-assisted groundwater bioremediation:Fundamentals, challenges and future perspectives[J]. Bioresource Technology, 196: 677-684. DOI:10.1016/j.biortech.2015.07.074 |
连玉倩, 何江涛, 梁雨, 等. 2018. 纳米乳化油强化硝酸盐反硝化产气变化研究[J]. 中国环境科学, 38(6): 2105-2115. DOI:10.3969/j.issn.1000-6923.2018.06.013 |
Liang S H, Kuo Y C, Chen S H, et al. 2013. Development of a slow polycolloid-releasing substrate (SPRS) biobarrier to remediate TCE-contaminated aquifers[J]. Journal of Hazardous Materials, 254: 107-115. |
Lindow N L, Borden R C. 2005. Anaerobic bioremediation of acid mine drainage using emulsified soybean oil[J]. Mine Water and the Environment, 24(4): 199-208. DOI:10.1007/s10230-005-0096-9 |
刘瑶.2017.胞外聚合物和废水中Cr(Ⅵ)的相互作用及对其厌氧生物还原的影响[D].大连: 大连理工大学 http://cdmd.cnki.com.cn/Article/CDMD-10141-1017822448.htm
|
吕英.2007.活性污泥胞外聚合物提取方法及其对重金属离子的吸附研究[D].天津: 天津大学 http://cdmd.cnki.com.cn/article/cdmd-10056-2008182575.htm
|
More T T, Yadav J S S, Yan S, et al. 2014. Extracellular polymeric substances of bacteria and their potential environmental applications[J]. Journal of Environmental Management, 144(1): 1-25. |
Newman W A, Pelle R C. 2006. Enhanced anaerobic bioremediation of chlorinated solvents utilizing vegetable oil emulsions[J]. Remediation Journal, 16: 109-122. |
Pham H L, Sechet P, Huang Z J.2018.A modified model for the prediction of bioclogging in saturated porous media[A]// Tran N H.Proceedings of the 4th congrès international de géotechnique-ouvrages-structures[C].Singapore: Springer Nature. 991-999
|
蒲晓芬, 胡涛, 周学东. 2005. 生物膜胞外聚合物的研究[J]. 国际口腔医学杂志, 32(5): 339-342. DOI:10.3969/j.issn.1673-5749.2005.05.004 |
生贺, 于锦秋, 刘登峰, 等. 2015. 乳化植物油强化地下水中Cr(Ⅵ)的生物地球化学还原研究[J]. 中国环境科学, 35(6): 1693-1699. DOI:10.3969/j.issn.1000-6923.2015.06.011 |
Sponza D T. 2003. Investigation of extracellular polymer substances (EPS) and physicochemical properties of different activated sludge flocs under steady-state conditions[J]. Enzyme and Microbial Technology, 32(3/4): 375-385. |
Tang G P, Wu W M, Watson D B, et al. 2013. U(Ⅵ) bioreduction with emulsified vegetable oil as the electron donor-microcosm tests and model development[J]. Environmental Science & Technology, 47(7): 3209-3217. |
唐远江, 雷帮星, 康冀川, 等. 2016. 缺陷假单胞菌HD13发酵过程中生物量和抗菌活性的变化[J]. 贵州农业科学, 44(4): 92-95, 99. DOI:10.3969/j.issn.1001-3601.2016.04.023 |
Thullner M. 2010. Comparison of bioclogging effects in saturated porous media within one- and two-dimensional flow systems[J]. Ecological Engineering, 36(2): 176-196. DOI:10.1016/j.ecoleng.2008.12.037 |
Thullner M, Schroth, Martin H, et al. 2004. Modeling of a microbial growth experiment with bioclogging in a two-dimensional saturated porous media flow field[J]. Journal of Contaminant Hydrology, 70(1/2): 37-62. |
Tsai T T, Liu J K, Chang Y M, et al. 2014. Application of polycolloid-releasing substrate to remediate trichloroethylene-contaminated groundwater:A pilot-scale study[J]. Journal of Hazardous Materials, 268: 92-101. DOI:10.1016/j.jhazmat.2014.01.004 |
Verardo E, Atteia O, Prommer H. 2017. Elucidating the fate of a mixed toluene, DHM, methanol, and i-propanol plume during in situ bioremediation[J]. Journal of Contaminant Hydrology, 201: 6-18. DOI:10.1016/j.jconhyd.2017.04.002 |
Vujasinovic S, Zaric J, Kaludjerovic D, et al. 2014. Anaerobic biodegradation of nitrate in groundwater sources ′Kljuc′ of the city of Pozarevac by the application of emulsified vegetable oil[J]. Zastita Materijala, 55: 69-75. DOI:10.5937/ZasMat1401069V |
Wang Y, Jin L Y, Deshusses M A, et al. 2013. The effects of various amendments on the biostimulation of perchlorate reduction in laboratory microcosm and flow through soil columns[J]. Chemical Engineering Journal, 232: 388-396. DOI:10.1016/j.cej.2013.07.060 |
Waston D B, Wu W M, Mehlhorn T, et al. 2013. In situ bioremediation of uranium with emulsified vegetable oil as the electron donor[J]. Environmental Science & Technology, 47(12): 6440-6448. |
魏复盛, 国家环境保护总局, 水和废水监测分析方法编委会. 2002. 水和废水监测分析方法(第四版)[M]. 北京: 中国环境科学出版社.
|
Wen C Y, Sheng H, Ren L M, et al. 2017. Study on the removal of hexavalent chromium from contaminated groundwater using emulsified vegetable oil[J]. Process Safety and Environmental Protection, 109: 599-608. DOI:10.1016/j.psep.2017.05.004 |
Xia L, Zheng X L, Shao H B, et al. 2016. Effects of bacterial cells and two types of extracellular polymers on bioclogging of sand columns[J]. Journal of Hydrology, 535: 293-300. DOI:10.1016/j.jhydrol.2016.01.075 |
杨靖, 叶淑君, 吴吉春. 2011. 生物膜对饱和多孔介质渗透性影响的实验和模型研究[J]. 环境科学, 32(5): 1364-1371. |
Yang S Y, Ngwenya B T, Butler I B, et al. 2013. Coupled interactions between metals and bacterial biofilms in porous media:implications for biofilm stability, fluid flow and metal transport[J]. Chemical Geology, 337-338: 20-29. DOI:10.1016/j.chemgeo.2012.11.005 |
Zhang P, Van Nostrand J D, He Z L, et al. 2015. A slow-release substrate stimulates groundwater microbial communities for long-term in-situ Cr(Ⅵ) reduction[J]. Environmental Science & Technology, 49(21): 12922-12931. |
Zhang Y S, Gillham R K. 2005. Effects of gas generation and precipitates on performance of Fe degrees PRBs[J]. Ground Water, 43(1): 113-121. |
赵阅坤.2016.乳化油碳源原位治理地下水中Cr(Ⅵ)静态批实验研究[D].北京: 中国地质大学(北京) http://cdmd.cnki.com.cn/Article/CDMD-11415-1016068094.htm
|
中华人民共和国农业部.2008.NY/T 1676-2008食用菌中粗多糖含量的测定[S].北京: 中国标准出版社
|