 2019, Vol. 39
2019, Vol. 39



2. 中国科学院生态环境研究中心, 中国科学院饮用水科学与技术重点实验室, 北京 100085;
3. 清华大学环境学院, 环境模拟与污染控制国家重点联合实验室, 水质与水生态研究中心, 北京 100084
2. Key Laboratory of Drinking Water Science and Technology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085;
3. Center for Water and Ecology, State Key Joint Laboratory of Environmental Simulation and Pollution Control, School of Environment, Tsinghua University, Beijing 100084
随着经济发展与人类活动加剧, 过量N、P的输入导致湖泊富营养化问题突出, 藻华频发, 生态功能退化(李娜等, 2018;马进, 2018;Wang et al., 2019;Zhao et al., 2019).富营养化引起的环境问题将可能成为中国未来可持续发展的主要问题(Wang et al., 2019).据统计, 20世纪70年代末, 我国富营养化湖泊仅占41%, 80年代后期发展到61%, 90年代后期高达77%(席北斗, 2013).其中, 具有代表性的太湖、滇池、巢湖和鄱阳湖已经受到不同程度的污染(Li et al., 2019;Wang et al., 2019), 并得到了相关专家学者的关注;与此同时, 程海湖逐渐进入了人们的视野, 当地环保部门也加强了对程海湖水质的监测, 但大都仅限于不同断面间的监测, 没有对整个程海湖进行系统性监测.
程海湖是全球最适宜螺旋藻天然生长的3个淡水湖泊之一, 地处云贵高原与滇西北高原衔接部位(100°38′~100°41′E, 26°17′~ 26°28′N), 为高原碱性湖泊, 隶属于云南省永胜县.湖体南北长19 km, 东西平均宽4.3 km, 水域面积约72.9 km2, 湖岸线长47.49 km, 最大水深32.2 m, 平均水深23.7 m, pH值9.03~9.42, 年平均水温17.8 ℃(董云仙等, 2012;杨世美等, 2018).自1960年以来, 程海湖水位逐年下降, 现已演变成一个封闭型湖泊, 其补给水源主要有湖面降水、地下水、流域内径流及外流域调水, 损耗主要是湖面蒸发、农田灌溉和工业用水(季克强, 2014;和继华, 2016).自1991年起, 程海湖南岸附近逐步发展起螺旋藻养殖企业, 之后进入稳定期, 导致湖泊流域输入水体中的营养物质发生了质和量的重大改变, 环境压力在持续加大(李松等, 2009).现阶段程海湖正面临着水位持续下降、水质恶化加快、水生态破坏严重等问题, 新时期亟待对程海湖富营养化状况进行评价并积极主动寻找对策保护程海湖.基于此, 本文对程海湖水质与浮游植物群落进行系统全面调研, 剖析浮游植物群落结构特征及进行驱动因子的识别, 并基于水质和藻类生物学两种方式评价程海湖的营养状态, 旨在为开展湖泊富营养化控制提供合理的参考意见, 并为程海湖水环境管理决策提供科学支撑.
2 材料与方法(Materials and methods) 2.1 样品采集与处理2018年7月—2019年1月对程海湖进行调查研究(逐月采样), 结合程海湖形态和入湖河流分布状况, 自南向北设置5个断面, 每个断面设3个采样点, 分别记为1#~15#, 共15个, 分布状况如图 1所示.采样期间, 首先利用GPS定位到达采样点, 用5 L有机玻璃采样器分别采集每个采样点表层(湖面以下0.5 m)和底层(湖底以上0.5 m)水样, 随即倒入经酸浸泡、清洗干净的聚乙烯采样瓶中.另各取1000 mL倒入棕色洁净聚乙烯瓶中, 并用15%鲁哥氏液(15 mL)固定, 后续用作藻类镜检(赵坤, 2018).收集完水样后, 当天冷藏即刻运往实验室于4 ℃环境保存, 并于短期内完成水质指标分析.
 |
| 图 1 采样点位置示意图 Fig. 1 Sampling sites schematic |
本次湖泊水质监测中理化指标测定参照《地表水和污水监测技术规范》 (HJ/T 91—2002)、《湖泊富营养化调查规范》和《水和废水监测分析方法》(第四版), 具体测定方法详见表 1.
| 表 1 理化指标测定方法 Table 1 Methods and standards for determination of physical |
综合营养状态指数法是利用水体基本理化指标进行富营养化评价的常用方法(金相灿, 1987).本文选取叶绿素a(Chla)、总氮(TN)、总磷(TP)、高锰酸盐指数(CODMn)和透明度(SD)5个参数来计算程海湖水体的综合营养状态指数(Li et al., 2019), 采用0~100数值进行营养状态分级, 表 2为评价标准(马千里等, 2014), 计算公式见式(1).
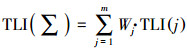
|
(1) |
| 表 2 综合营养状态指数评价标准 Table 2 Assessment standard of eutrophication by comprehensive trophic state index |
式中, TLI(∑)是综合营养状态指数, Wj是第j种参数的营养状态指数的相关权重, TLI(j)代表第j种参数的营养状态指数.
2.2.2 藻类生物学法浮游植物是湖泊水生生态系统中的重要部分, 当水环境有所变化时, 水中藻类的密度、种类与结构组成也会随之发生相应的变化, 能够在一定程度上反映湖泊的水质情况, 并可用于预测水生生态系统的变化趋势.浮游藻类的细胞密度可以在一定程度上反映湖泊的营养程度, 一般藻密度小于30×104个·L-1的湖泊为贫营养型, 在30×104~100×104个·L-1之间的为中营养型, 大于100×104个·L-1的为富营养型(Zhang et al., 2018);浮游藻类的细胞密度也可以用来评价水华程度, 其标准详见表 3.另外, 藻类的物种多样性可以说明不同环境条件下藻类个体的分布丰度和水体污染程度.本研究选取应用较为广泛的Shannon-Wiener多样性指数H′(式(2))和Pielou均匀度指数J(式(3))(孙军等, 2004), 对程海湖各个采样点的水质进行评价, 藻类群落多样性和均匀度指数对水环境状况的指示作用评价标准参见表 4(Shanthala et al., 2009).
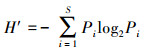
|
(2) |
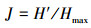
|
(3) |
| 表 3 中国环境监测总站制定的藻类水华特征评价方法和分级标准 Table 3 Assessment methods and grading standards for algal bloom characteristics formulated by China National Environmental Monitoring Centre |
| 表 4 浮游植物水质生物学评价标准 Table 4 Bioassessment standard for phytoplankton in water quality |
式中, Pi=ni/N;Hmax=log2S, 为最大多样性指数;ni为第i种的个体数量(ind.·m-3);N为某站总生物数量(ind.·m-3);S为出现生物总种数.
3 结果与讨论(Results and discussion) 3.1 水质状况分析表 5为程海湖水质基本指标结果, 夏季(7月)与冬季(1月)水温差别较大, pH值范围为9.21~9.51, 均值为9.36, 依然维持碱性湖泊的性质, 时空差异较小. Chla值的范围为10.23~73.61 μg·L-1, 均值为31.09 μg·L-1, 已经具备水华现象的指征. CODMn常被用来作为湖泊水体有机污染的评价指标, 本次调查中程海湖CODMn的变化范围为1.14~8.41 mg·L-1, 均值为5.11 mg·L-1, 较丽江市环境监测站历史监测结果有小幅度提高.表 6列出的是2018年期间夏、冬两个季节程海湖营养盐变化状况, TP浓度范围为0.03~0.19 mg·L-1, TN为0.38~3.08 mg·L-1, 平均值分别是0.07、1.26 mg·L-1, 最高值分别是最低值的4.75、6.45倍, 具有发生水华的营养盐基础.参照《地表水环境质量标准》(GB 3838—2002), 监测期间水质状况已经无法达到Ⅲ类水保护目标, 除pH外, 主要超标项目是TN和TP, 已经达到富营养水平.在夏、冬季节交替的过程中, TN有较大幅度上升, TP基本保持不变.夏、冬两季表层总磷浓度均略低于底层, 结合2018年实测底泥沉积物中TP均值分别为0.93 g·kg-1, 说明可能与水底缺氧引起的底层沉积物中磷释放有关(Bai et al., 2019).
| 表 5 2018年夏、冬两季程海湖水质特征 Table 5 Characteristics of water quality in Chenghai Lake in summer and winter, 2018 |
| 表 6 2018年夏、冬两季程海湖营养盐变化情况 Table 6 Changes of nutrient in Chenghai Lake in summer and winter, 2018 |
浮游藻类的生长高度依赖于水文条件和理化参数(Abd El-Hady et al., 2016;Jiang et al., 2019), 而氮磷比(TN/TP)是表征水体营养盐结构的主要指标, 也是水体中浮游植物营养结构特点的一个重要参考指标, 对于衡量藻类暴发性生长具有十分重要的意义(刘淼等, 2017;Tong et al., 2018;Li et al., 2019).有研究提出, 当湖水TN和TP浓度比在10:1~25:1时, 藻类生长与氮、磷浓度存在线性相关关系(樊鑫, 2018).夏季时期程海湖水质情况正处于该区间, 属于有利于藻类生长的营养条件.还有研究提出, TN/TP>10为磷限制性状态, TN/TP<7为氮限制性状态(Xie et al., 2003;单保庆等, 2012), 据此则可初步判定在本次调研期间程海湖基本处于磷限制性状态.与2015年的水质状况(TN、TP、NH4+-N、CODMn均值为0.05、0.69、0.11、3.70 mg·L-1)相比, 当前程海湖的水质已达到富营养化水平, 未来可能会引发水中藻类大量繁殖.
3.2 浮游植物群落结构特征本研究对程海湖水域采样过程中, 15个采样点表、底层全覆盖, 保证了调研数据的完整性, 可以最大限度地反映出程海湖藻类构成的真实可靠性.本次共鉴定出7门78种浮游藻类, 从种类上看, 种类数目较多的依次是绿藻门、硅藻门和蓝藻门, 其中, 绿藻门有33种, 占种类总数的42.3%;硅藻门有19种, 占种类总数的24.4%;蓝藻门有14种, 占种类总数的17.9%.图 2为2018年程海浮游藻类的种类组成分布图, 与表 7中的历史数据(董云仙等, 2012)进行比较, 发现藻类种群结构中硅藻比重下降, 而蓝、绿藻比重有所上升, 表明程海湖富营养化进一步发展, 更加趋向于富营养化的蓝藻种群结构特征, 进一步印证了其富营养化的水质现状.
 |
| 图 2 2018年程海湖浮游藻类的种类组成 Fig. 2 Species composition of planktonic in Chenghai Lake, 2018 |
| 表 7 程海湖1985年与2010年藻类组成情况 Table 7 Comparison of algal community composition between 1985 and 2010 in Chenghai Lake |
从生物量上来看, 优势种主要为蓝藻门的柱孢鱼腥藻、伪鱼腥藻、拟柱孢藻、圆柱鱼腥藻之间相互演替, 检出比例高达50%以上, 多数为α-中污和β-中污指示种(洪尚群等, 2012), 一小部分甚至能够达到重度污染等级, 其中, 蓝藻数量呈现暖季多、冷季少的规律, 硅藻则相反, 这主要是因为蓝藻色素能耐高温, 且适于生长在有机质丰富的水体中(刘霞, 2012).各个采样点的藻类群落和优势藻种月度之间存在一定变化, 可能与藻类生长受到温度与光照变化的影响较大有关.这与文献(董云仙等, 2012)记载中程海湖优势藻种呈现季节变换规律类似, 虽然优势藻种有变化, 但优势藻种依旧属于蓝藻门.
3.3 浮游植物分布与环境因子的关系为探究程海湖浮游植物群落分布的影响因素, 以2018年7月—2019年1月各月采样实测数据为基础, 选取可能性较大的环境因子进行Pearson相关性分析, 结果(表 8)表明, 藻细胞密度与Chla呈显著正相关(p < 0.01), 这一点印证了Chla是表征水体中浮游植物数量的重要参数, 并且与二者呈正相关的环境因子具有较好的一致性.此外, 藻细胞密度还与DO、pH、ORP、TN、TOC、PO43--P、TDP、CODMn呈显著正相关.在藻细胞密度较高期间, 光合作用增强, 水体中的溶解氧增大, CO2浓度降低, 从而引起pH升高.此外, 也能看出浮游植物的生长受到表征有机污染的CODMn的影响较大.因此, 如果污染负荷得不到有效控制, 将会增加藻类水华的风险.
| 表 8 程海湖浮游植物群落与环境因子的Pearson相关性分析 Table 8 Pearson correlation analysis of phytoplankton community and environmental factors in Chenghai Lake |
富营养化是藻类在水生系统中营养物质的富集, 特别是氮和磷会导致藻类加速生长.本次评价依据中国环境监测总站推荐的“湖泊水库富营养化评价方法及分级技术规定”, 分析了所选定的程海湖15个采样点的营养状况.图 3为2018年7月—2019年1月期间各月的综合营养状态指数均值, 可以看出程海湖整体已经达到轻度富营养化水平, 隶属轻度污染范围.部分样点在某个时间段综合营养状态指数能够达到60, 综合营养指数的最大值几乎能够达到中度富营养化水平.这种情况可能与当地螺旋藻养殖企业废水泄漏、畜禽养殖、污水处理设施不健全及农业面源污染无法得到有效控制有一定关系.此外, 结合图 4中近十余年程海湖富营养化趋势可以看出, 富营养化程度在加重, 加之本身含有较高的氮、磷营养盐且还在不断积累, 这将会为藻类繁殖提供条件, 说明程海湖营养状况已经不容乐观.
 |
| 图 3 综合营养状态指数月均值 Fig. 3 Monthly mean of comprehensive trophic state index |
 |
| 图 4 2006—2017年程海湖综合营养状态指数 Fig. 4 Comprehensive trophic status index in Chenghai Lake from 2006 to 2017 |
基于理化指标的综合营养状态指数法评价湖泊的富营养化程度具有瞬时性, 不能较好地反映湖泊的健康状况, 而浮游植物作为湖泊生态系统的重要组成部分, 敏感性较强, 其群落结构、藻细胞密度及优势种对水体的营养状态都有直接或者间接的指示作用(Cupertino et al., 2019), 因此, 从藻类生物学角度来识别湖泊的营养状况能起到相辅相成的作用.藻类的细胞密度作为水生生态系统功能和水质评价的重要参数之一(Zhang et al., 2019), 在一定程度上也能反映水体的污染程度.图 5是2018年7月—2019年1月半年期间程海湖表层和底层的藻细胞密度均值(12月份样品保存失误, 故暂缺), 监测期间表层藻密度变化范围是2.07×106~1.65×107个·L-1, 均值为6.00×106个·L-1;底层为1.54×106~ 5.89×106个·L-1, 均值为3.47×106个·L-1.整体上表层藻细胞密度高于底层, 以丰水期(7月)较高、枯水期(1月)较低为主要规律, 受温度、光照及营养盐的影响较大.与董云仙等(2012)于2009年10月—2010年9月的调查数据(3.90×106~5.14×108个·L-1, 年平均4.85×107个·L-1)相比有所改善, 但数量依然庞大.参照中国环境监测总站制定的藻类水华特征评价方法和分级标准(表 3), 当前程海湖大部分时间处于轻微水华范围, 7、8月达到轻度水华水平, 可判断程海湖水华风险介于初级预警和中级预警之间(杨刚, 2009), 水华发生的可能性较大.
 |
| 图 5 2018年7月—2019年1月藻细胞密度变化曲线 Fig. 5 The change curve of algal cell density from July 2018 to January 2019 |
水体的浮游植物中, 各大门类的藻类能够适应并生存在不同营养类型的湖泊中, 从而在一定程度上指示湖泊的营养状况.一般以金藻、黄藻类为主的属于贫营养型湖泊, 以甲藻、硅藻、隐藻为主的属于中营养型, 而富营养型湖泊常以蓝藻和绿藻为优势种(Zhang et al., 2019).图 6为程海湖夏、冬两季节不同采样点浮游藻类的相对丰度, 可以看出, 程海湖藻类主要以蓝藻门、绿藻门、甲藻门和硅藻门为主, 丰度占比能够达到95.6%.虽然各个采样点不同藻门所占比例有所不同, 但整体上还是以蓝藻为优势种, 并且指示较清洁水体的金藻仅在7号采样点有极少量出现, 说明程海湖已经达到富营养湖泊水平.
 |
| 图 6 不同采样点浮游藻类相对丰度 (a.2018年7月, b.2019年1月) Fig. 6 Relative abundance of planktonic algae at different sampling sites (a.July 2018, b.January 2019) |
浮游植物多样性指数和均匀度指数也可以较好地反映水质变化状况, 水中检出的藻类种数越多, 并且各个藻种的分布均匀性越高, 说明该水体中浮游植物群落的稳定性越高, 水环境质量也越高.图 7中2018年7月和2019年1月的两次监测结果显示, 程海湖Shannon-Wiener多样性指数大多介于1~3, 整体上夏季(7月)低于冬季(1月), 说明冬季较夏季浮游植物群落结构稳定, 水体污染程度较低, 这与7月藻密度值高于1月的水质污染评价结果相对应.各采样点Pielou均匀度指数夏季(7月)变幅为0.18~0.62, 均值为0.33, 冬季(1月)变幅为0.42~0.72, 均值为0.61, 与Shannon-Wiener多样性指数评价结果保持一致, 大部分评价为α中度污染和β中度污染, 个别采样点(4、9和15号)在夏季7月甚至达到了重度污染等级, 与理化指标评价结果具有较好的一致性.
 |
| 图 7 多样性指数与均匀度指数分布 (a.Shannon-Wiener多样性指数, b.Pielou均匀度指数) Fig. 7 Diversity index and evenness index distribution (a.Shannon-Wiener diversity index, b.Pielou evenness index) |
1) 根据最新监测结果并结合历史数据分析, 程海湖水质状况呈下降趋势, 总体水质评价为Ⅳ类水, 主要污染因子为氮、磷, 总磷(TP)浓度为0.03~0.19 mg·L-1, 总氮(TN)浓度为0.38~3.08 mg·L-1, 已超过常规湖泊的水华暴发阈值范围, 在监测期间各采样点的综合营养状态指数(TLI)在49.3~54.4之间波动, Chla值的范围为10.23~73.61 μg·L-1, 基本处于轻度富营养化水平.
2) 调查期间, 程海湖藻类共计7门78种, 藻密度范围为1.54×106~1.65×107个·L-1.水华风险方面, 整体上处于轻微水华水平, 部分时间发生轻度水华, 藻类水华风险在不断加大.Shannon-Wiener多样性指数大多介于1~3, Pielou均匀度指数介于0.18~0.72, 部分时间段藻类多样性偏低, 且数量上以蓝藻门和绿藻门为主, 受温度和水体富营养化程度影响较为明显, 优势种主要有柱孢鱼腥藻、伪鱼腥藻、拟柱孢藻和圆柱鱼腥藻, 指示为典型的富营养化特征, 与理化指标评价结果具有较好的一致性.
3) 整体上程海湖浮游植物种类组成和水环境特征都有所变化, 藻密度与Chla、DO、pH、ORP、TN、TOC、PO43--P、TDP、CODMn均呈显著正相关.进一步研究程海湖浮游植物群落结构变化状况很有意义, 只有通过长期科学的监测与防治, 加之有效的生态修复措施, 才能逐步缓解程海湖的富营养化现状, 从源头上降低水华暴发的风险.
致谢(Acknowledge): 特别感谢丽江环境监测站和永胜县环保局提供的数据材料及程海管理局相关同志在调研采样期间给予的大力支持与帮助.
Abd El-Hady H H, Fathey S A, Ali G H, et al. 2016. Biochemical profile of phytoplankton and its nutritional aspects in some khors of Lake Nasser, Egypt[J]. Egyptian Journal of Basic and Applied Sciences, 3(2): 187-193. DOI:10.1016/j.ejbas.2016.03.002 |
Bai Y, Zeng Y, Nie B, et al. 2019. Hydrodynamic disturbance on phosphorus release across the sediment-water interface in Xuanwu Lake, China[J]. Water Science and Technology-Water Supply, 19(3): 735-742. DOI:10.2166/ws.2018.115 |
Cupertino A, Gücker B, Von Rückert G, et al. 2019. Phytoplankton assemblage composition as an environmental indicator in routine lentic monitoring:Taxonomic versus functional groups[J]. Ecological Indicators, 101: 522-532. DOI:10.1016/j.ecolind.2019.01.054 |
单保庆, 菅宇翔, 唐文忠, 等. 2012. 北运河下游典型河网区水体中氮磷分布与富营养化评价[J]. 环境科学, 33(2): 352-358. |
董云仙, 洪雪花, 谭志卫. 2012. 高原深水湖泊程海中氮元素时空分布特征[J]. 环境科学与技术, 35(9): 173-178. DOI:10.3969/j.issn.1003-6504.2012.09.036 |
董云仙, 谭志卫, 朱翔, 等. 2012. 程海藻类植物种群结构和数量的周年变化特征[J]. 生态环境学报, 1(7): 1289-1295. |
El-Serehy H A, Abdallah H S, Al-Misned F A, et al. 2018. Assessing water quality and classifying trophic status for scientifically based managing the water resources of the Lake Timsah, the lake with salinity stratification along the Suez Canal[J]. Saudi Journal of Biological Sciences, 25(7): 1247-1256. DOI:10.1016/j.sjbs.2018.05.022 |
樊鑫.2018.长江口及邻近海域沉积硅藻记录及富营养化历史重建[D].北京: 中国科学院大学(中国科学院海洋研究所)
|
和继华. 2016. 程海湖补水可行性研究[J]. 环境科学导刊, 35(1): 35-38. DOI:10.3969/j.issn.1673-9655.2016.01.008 |
洪尚群, 梁川, 张瀚, 等. 2012. 云南松花坝水源区水质藻类生物学评价与影响藻类生长的因素研究[J]. 水力发电学报, 31(2): 152-158. |
金相灿. 1987. 湖泊富营养化调查规范[M]. 北京: 中国环境科学出版社.
|
Jiang Z, Du P, Liu J, et al. 2019. Phytoplankton biomass and size structure in Xiangshan Bay, China:Current state and historical comparison under accelerated eutrophication and warming[J]. Marine Pollution Bulletin, 142: 119-128. DOI:10.1016/j.marpolbul.2019.03.013 |
季克强.2014.程海湖水位的历史演变及影响因素分析[A].中国科学技术协会.湖泊保护与生态文明建设-第四届中国湖泊论坛论文集[C].中国科学技术协会: 安徽省科学技术协会学会部
|
Li C, Feng W, Chen H, et al. 2019. Temporal variation in zooplankton and phytoplankton community species composition and the affecting factors in Lake Taihu-a large freshwater lake in China[J]. Environmental Pollution, 245: 1050-1057. DOI:10.1016/j.envpol.2018.11.007 |
李娜, 黎佳茜, 李国文, 等. 2018. 中国典型湖泊富营养化现状与区域性差异分析[J]. 水生生物学报, 42(4): 854-864. |
李松, 王钟. 2009. 永胜县程海湖螺旋藻产业发展与环保问题调查[J]. 时代金融, (12): 158-159. DOI:10.3969/j.issn.1672-8661.2009.12.090 |
刘淼, 陈开宁, 孙淑雲, 等. 2017. 大溪水库水质变化趋势及污染成因解析[J]. 环境科学学报, 37(7): 2514-2523. |
刘霞.2012.太湖蓝藻水华中长期动态及其与相关环境因子的研究[D].武汉: 华中科技大学
|
马进.2018.富营养水体生态修复过程中氮、磷营养盐迁移转化研究[D].上海: 上海海洋大学
|
马千里, 胡芳, 虢清伟, 等. 2014. 六都寨水库夏季富营养化状况与浮游植物分布特征研究[J]. 环境科学学报, 34(6): 1497-1504. |
Shanthala M, Hosnani S P, Hosetti B B. 2009. Diversity of phytoplanktons in a waste stabilization pond at Shimoga Town, Karnataka State, India[J]. Environmental Monitoring & Assessment, 151: 437-443. |
孙军, 刘东艳. 2004. 多样性指数在海洋浮游植物研究中的应用[J]. 海洋学报(中文版), (1): 62-75. DOI:10.3321/j.issn:0253-4193.2004.01.007 |
Tong Y, Qiao Z, Wang X, et al. 2018. Chemosphere Human activities altered water N:P ratios in the populated regions of China[J]. Chemosphere, 210: 1070-1081. DOI:10.1016/j.chemosphere.2018.07.108 |
Wang J H, Yang C, He L Q S, et al. 2019. Meteorological factors and water quality changes of Plateau Lake Dianchi in China (1990-2015) and their joint in fluences on cyanobacterial blooms[J]. Science of the Total Environment, 665: 406-418. DOI:10.1016/j.scitotenv.2019.02.010 |
Wang J, Fu Z, Qiao H, et al. 2019. Assessment of eutrophication and water quality in the estuarine area of Lake Wuli, Lake Taihu, China[J]. Science of the Total Environment, 650: 1392-1402. DOI:10.1016/j.scitotenv.2018.09.137 |
席北斗. 2013. 湖泊营养物标准方法学及案例研究[M]. 北京: 科学出版社.
|
Xie L Q, Xie P, LI S X, et al. 2003. The low TN:TP ratio, a cause or a result of Microcystis blooms?[J]. Water Research, 37: 2073-2080. DOI:10.1016/S0043-1354(02)00532-8 |
杨刚.2009.洪泽湖藻类监测与"水华"预警模式研究[A]//中国环境科学学会2009年学术年会论文集(第一卷)[C].北京: 中国环境科学学会
|
杨世美, 汪涛, 王泽平, 等. 2018. 程海湖水生态承载力变化研究[J]. 水利水电快报, 39(2): 39-44. |
Zhang C, Yao X, Chen Y, et al. 2019. Variations in the phytoplankton community due to dust additions in eutrophication, LNLC and HNLC oceanic zones[J]. Science of the Total Environment, 669: 282-293. DOI:10.1016/j.scitotenv.2019.02.068 |
Zhang M, Shi X, Yang Z, et al. 2018. Long-term dynamics and drivers of phytoplankton biomass in eutrophic Lake Taihu[J]. Science of the Total Environment, 645: 876-886. DOI:10.1016/j.scitotenv.2018.07.220 |
Zhao C S, Shao N F, Yang S T, et al. 2019a. Integrated assessment of ecosystem health using multiple indicator species[J]. Ecological Engineering, 130: 157-168. DOI:10.1016/j.ecoleng.2019.02.016 |
Zhao C S, Shao N F, Yang S T, et al. 2019b. Quantitative assessment of the effects of human activities on phytoplankton communities in lakes and reservoirs[J]. Science of the Total Environment, 665: 213-225. DOI:10.1016/j.scitotenv.2019.02.117 |
赵坤.2018.淮河流域浮游动物分布格局及群落形成机制[D].武汉: 华东师范大学
|
张佐. 2013. 永胜县筑起微藻之都[J]. 中国农业信息, (1): 32-33. DOI:10.3969/j.issn.1672-0423.2013.01.013 |