 2019, Vol. 39
2019, Vol. 39



2. 湿润亚热带生态-地理过程教育部重点实验室, 福州 350007;
3. 福建师范大学亚热带湿地研究中心, 福州 350007
2. Key Laboratory of Humid Sub-tropical Eco-geographical Process of Ministry of Education, Fuzhou 350007;
3. Research Centre of Wetlands in Subtropical Region, Fujian Normal University, Fuzhou 350007
甲烷(CH4)作为一种重要的温室气体, 具有很强的红外吸收能力和高效的单分子增温潜势(Sun et al., 2013), 其在大气中的浓度已从工业革命前的0.722 μL · L-1增加到2011年的1.803 μL · L-1, 百年内的增温潜势为CO2的28倍, 辐照强度占大气层中温室气体总辐射强度的18%, 强烈影响着全球热平衡(IPCC, 2013).自然湿地作为大气中CH4的主要排放源, 每年向大气排放177~284 Tg CH4, 占全球CH4总排放量的26%~42%, 在全球碳循环中具有十分重要的作用(IPCC, 2013).河口湿地作为自然湿地重要组成部分, 是大气环境中CH4不可忽视的自然源.
在全球气候变化和人类活动双重作用下, 滨海及河口地区, 由于海平面的不断上升, 以及地表水和地下水的过量开采, 加之地势低平, 海水入侵强度日趋增加(Loáiciga et al., 2012; Neubauer, 2013), 河口湿地正在或将面临盐水入侵的影响.盐度的改变将深刻影响河口湿地水-土环境, 植物群落初级生产力和有机物积累速率会因此降低, 还会改变微生物作用下的有机物矿化速率和途径等(Weston et al., 2006; Neubauer, 2013), 进而影响湿地生态系统的结构和功能(Kiehn et al., 2013).关于盐度增加对于河口湿地CH4产生与排放的影响, 主要表现为盐水入侵在一定程度上会抑制CH4的产生与排放(Pivni-ková et al., 2010;Chambers et al., 2011);但也有研究表明, 盐水入侵有可能不足以对CH4产生抑制作用(Konnerup et al., 2014), 甚至促进CH4的产生(Weston et al., 2006), 或影响不显著(曾从盛等, 2008; Krauss et al., 2011;姜欢欢等, 2012).
湿地土壤产生的CH4, 一部分通过气泡、扩散和植物传输3种方式进入到大气中, 而另一部分则在植物根系周围或水气界面被氧化, 还有相当一部分CH4溶解在土壤孔隙水中, 长期或短期储存在土壤中.有研究发现, 土壤表层孔隙水中CH4浓度能够解释CH4排放通量的91%(Ding et al., 2005);在滨海沼泽湿地中, CH4排放通量和表层5 cm孔隙水中甲烷浓度的相关系数也可达到0.45~0.73(Ding et al., 2010).
海平面的上升可能会造成河口区淡水感潮湿地盐度的长期增加, 而台风等风暴潮的发生则可能会造成短期的盐水上溯, 使得河口区淡水无感潮湿地盐度短时增加.本研究以闽江河口区塔礁洲淡水感潮短叶茳芏沼泽湿地及其比邻的无植被覆盖光滩湿地为研究对象, 通过构建中型生态系统的方法, 探讨恒定和波动盐度增加对河口淡水感潮湿地间隙水溶解性CH4浓度及间隙水化学特征的影响, 以期为今后准确估测长期和短期盐水入侵对我国亚热带河口感潮湿地甲烷动态的影响提供科学数据, 并更深入地揭示不同盐水入侵情景下河口淡水感潮湿地间隙水甲烷动态特征.
2 材料与方法(Materials and methods) 2.1 研究样地塔礁洲位于闽江河口区中部(119°18′29″~119°22′04″E, 25°58′03″~25°57′37″N), 面积约878 hm2, 属于淡水洲滩, 洲中心已开发居民点和农田, 洲四周主要由沙滩和泥滩组成, 短叶茳芏等湿地植物分布于泥滩湿地边缘及地势低洼区(刘剑秋等, 2010).本研究采样点位于塔礁洲南岸(图 1).塔礁洲南岸分布有较大面积的短叶茳芏沼泽湿地及其附近的无植被覆盖光滩(以下简称为光滩)(图 2), 采样点土壤理化性质见表 1.
 |
| 图 1 研究区与采样点位置图 Fig. 1 Location of study area and sampling site |
 |
| 图 2 采样点实景 Fig. 2 Picture of sampling sites |
| 表 1 采样点土壤理化性质 Table 1 Soil physical and chemical properties of the sampling site |
含有土壤+植物系统的中型实验生态系(Mesocosm)既保持了原自然生态系统的完整性, 又便于控制影响因子, 在揭示某一生态因子对生态系统过程影响的研究中发挥着重要作用, 常用于全球变化对湿地生态系统过程和功能影响的控制性实验研究, 本研究基于中型实验生态系开展相关控制实验研究.2016年11月采集塔礁洲南岸短叶茳芏沼泽湿地植物-土芯连续体及其附近光滩土芯各9根(土芯深度30 cm, 直径25 cm), 构建中型实验生态系.对于短叶茳芏沼泽湿地, 土芯和生长的短叶茳芏整体采集, 放置在容器内, 紧密贴合容器内壁, 运回校园构建中型实验生态系统, 开展模拟恒定和波动盐度增加对河口淡水感潮间隙水溶解性CH4浓度影响的研究.为避免降水对实验带来的干扰, 对中型生态系统进行遮雨防护.中型实验生态系统保持长期淹水状态(水淹深度5~6 cm).
取样样地为典型的淡水感潮沼泽湿地, 故本研究设置了恒定淡水、恒定盐度增加(主要模拟持续的海平面上升造成的淡水感潮湿地“永久性的”盐度增加), 以及波动盐度增加(主要模拟风暴潮等引起的淡水感潮湿地“短期的”盐度增加)3个处理, 波动盐度增加设为每个实验周期的前3周添加淡水(0‰), 第4周添加咸水(10‰), 每个处理3个重复.对于短叶茳芏中型实验生态系统, 3个处理分别记为恒定淡水(Fw-V)、恒定盐度(Sw-V)和波动盐度(Fl-V);对于光滩中型实验生态系, 3个处理分别记为恒定淡水(Fw-M)、恒定盐度(Sw-M)和波动盐度(Fl-M).实验于2016年12月开始, 至2017年10月结束, 每4周为1个周期, 恒定淡水处理一直添加盐度为0‰的当地淡水;恒定盐度处理一直添加利用海盐与蒸馏水按比例配置的10‰盐度的人造盐水;波动盐度处理的盐度变化设定为每个实验周期的前3周添加当地淡水, 第4周添加10‰的人造盐水.
2.3 中型生态系统间隙水采集与测定利用间隙水采样器(Porewater samplers, Rhizon samplers)(Roos et al., 2007; Geurts et al., 2010; 罗敏等, 2016)采集中型生态系统土壤表层以下5 cm处的间隙水.2017年1—9月, 每4周采集1次.未淹水前提前将采水管透过容器壁水平插入土壤表层以下5 cm深处, 为防止间隙水从容器侧壁渗漏及土壤与外界空气接触, 用防水胶将软管口周围密封, 管口安装一个三通阀, 在不采集水样时关闭;采集水样时先将注射器针筒抽至真空状态, 再与软管口连接, 确保连接紧密后打开三通阀进行水样采集.
间隙水溶解性CH4浓度测定采用顶空平衡-气相色谱法(Kazunori et al., 2010).将注射器中的间隙水(10 mL)注射到已抽真空的20 mL顶空瓶中, 再用气密性注射器向顶空瓶中注入10 mL高纯N2(>99.999%).室温下将样品瓶剧烈振荡30 min, 静置30 min, 待瓶内气-液两相达到平衡, 抽取3.5 mL顶空气体注入气相色谱仪(GC-2010, Shimadzu)测定CH4浓度, 根据以下公式计算间隙水溶解性CH4浓度:
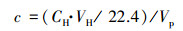
|
(1) |
式中, c为溶解性CH4浓度(μmol · L-1), CH为顶空瓶上部空间CH4浓度(μL · L-1), VH为顶空瓶上部空间体积(mL), VP为顶空瓶间隙水体积(mL).
剩余的间隙水经0.45 μm滤膜过滤后置于50 mL聚乙烯瓶保存, 并进行间隙水其他理化指标的测定.pH采用pH计(STARTER 3100)测定;盐度采用盐度计(Eutech Instruments SALT 6+, USA)测定;NH4+-N、NO3--N及TN浓度用流动注射分析仪(SKALAR San++, Netherlands)测定, 仪器检测限为0.005 mg · L-1;Cl-、SO42-浓度采用ICS-2100离子色谱仪(DIONEX, USA)测定, 检测限分别为0.002 mg · L-1和0.009 mg · L-1.
2.4 数据处理与分析运用Microsoft Excel 2003、OriginPro 8.0、和SPSS 19.0统计分析软件对数据进行整理与分析.平均值与标准误采用Microsoft Excel 2003软件计算;采用OriginPro 8.0软件作图;采用SPSS 19.0统计分析软件中的皮尔逊相关系数(Pearson′s correlation coefficient)进行相关分析, 并对数据进行线性回归分析, 采用重复测量方差分析及LSD检验进行方差分析, 影响因素分析采用单因变量多因素方差分析.
3 结果(Results) 3.1 短叶茳芏湿地及光滩土壤基本理化性质比较模拟实验开展前, 测定短叶茳芏湿地及光滩土壤基本理化性质并进行比较, 结果表明, 短叶茳芏湿地土壤总碳、总氮、Cl-、NH4+-N含量均显著高于光滩(p < 0.05), pH和NO3--N含量显著低于光滩(p < 0.05), SO42-含量差异不显著(p>0.05).
3.2 气温月动态特征图 3为2017年1—9月的气象因子变化特征.其中, 2月气温最低, 仅为7.69 ℃, 从3月开始突然升高, 达到26.83 ℃, 之后呈波动上升趋势, 并在9月达到最高值34.63 ℃, 总体表现为冬季(1—2月)气温低, 春季(3—5月)开始气温迅速上升, 夏、秋季持续高温(6—9月), 整个研究期间气温均值为25.72 ℃.
 |
| 图 3 研究区气温随时间的变化 Fig. 3 Temporal variation of temperature in study area |
间隙水溶解性CH4浓度具有明显的季节变化(图 4), 短叶茳芏湿地和光滩湿地3个处理间隙水溶解性CH4浓度最低值均出现冬季的1月, 最高值均出现在夏季的5—6月.短叶茳芏湿地Fw-V处理最低值和最高值分别为(0.12±0.02)和(65.18±3.50) μmol · L-1, 平均值为(23.41±5.29) μmol · L-1;Fl-V处理最低值和最高值分别为(0.14±0.01)和(19.49±3.60) μmol · L-1, 平均值为(4.20±1.22) μmol · L-1;Sw-V处理最低值和最高值分别为(0.11±0.01)和(2.09±0.99) μmol · L-1, 平均值为(0.61±0.15) μmol · L-1.光滩湿地Fw-M处理最低值和最高值分别为(0.17±0.05)和(80.35±4.43) μmol · L-1, 平均值为(33.93±6.09) μmol · L-1;Fl-M处理最低值和最高值分别为(0.39±0.01)和(49.62±14.92) μmol · L-1, 平均值为(14.20±3.50) μmol · L-1;Sw-M处理最低值和最高值分别为(0.11±0.01)和(1.23±1.08) μmol · L-1, 平均值为(0.48±0.12) μmol · L-1.
 |
| 图 4 恒定和波动盐度处理下间隙水溶解性CH4浓度的时间变化 Fig. 4 Temporal variations of porewater dissolved CH4 concentration under the constant and fluctuating salinity treatments |
多因素方差分析表明, 盐度恒定和波动变化、有无植被覆盖及气温均显著影响湿地间隙水溶解性CH4浓度(表 2), 光滩间隙水溶解性CH4浓度显著大于短叶茳芏湿地(p <0.05).不论是恒定盐度还是波动盐度增加处理, 均显著降低湿地土壤间隙水溶解性CH4浓度(p < 0.05), 短叶茳芏湿地中, 与恒定淡水处理相比, 波动盐度和恒定咸水处理下的间隙水溶解性CH4浓度年均值分别下降了82.1%和97.4%;光滩湿地中, 相对于淡水对照处理, 波动盐度和恒定咸水处理下的间隙水溶解性CH4浓度年均值分别下降了58.2%和98.6%.
| 表 2 间隙水溶解性CH4浓度多因素方差分析结果 Table 2 Results of multi-factor variance analysis for porewater dissolved CH4 concentration |
间隙水pH值月动态见图 5.短叶茳芏湿地中, Fw-V处理最低值和最高值分别出现在2月和6月, 年平均值为(6.41±0.13);Fl-V处理最低值和最高值分别出现在3月和1月, 年平均值为(5.28±0.16);Sw-V处理最低值和最高值分别出现在3月和8月, 年平均值为(4.91±0.16).光滩中, Fw-M处理最低值和最高值分别出现在1月和6月, 年平均值为(7.34±0.10);Fl-M处理最低值和最高值分别出现在5月和9月, 年平均值为(6.07±0.15);Sw-M处理最低值和最高值分别出现在8月和2月, 年平均值为(6.08±0.07).
 |
| 图 5 恒定和波动盐度处理下间隙水pH值的时间变化 Fig. 5 Temporal variation of porewater pH under constant and fluctuating salinity treatments |
多因素方差分析表明, 盐度、植物、时间均对河口湿地间隙水pH值有显著影响(表 3), 光滩湿地间隙水pH值显著大于短叶茳芏湿地(p < 0.05).恒定咸水及波动盐度处理均显著降低湿地土壤间隙水pH值(p < 0.05), 短叶茳芏湿地中, 与恒定淡水处理相比, 波动盐度和恒定咸水处理下的间隙水pH值年均值分别下降17.63%和23.40%;光滩湿地中, 相对于恒定淡水处理, 波动盐度和恒定咸水处理下的间隙水pH值年均值分别下降了17.30%和17.17%.
| 表 3 间隙水理化因子多因素方差分析结果 Table 3 Results of multi-factor variance analysis for porewater physical and chemical factors |
间隙水Cl-浓度月动态见图 6.短叶茳芏湿地中, Fw-V处理最低值和最高值分别出现在4月和2月, 年平均值为(41.69±4.01) mg · L-1;Fl-V处理最低值和最高值分别出现在1月和9月, 年平均值为(6301.64±535.40) mg · L-1;Sw-V处理最低值和最高值分别出现在1月和5月, 年平均值为(16741.93±960.93) mg · L-1.光滩中, Fw-M处理最低值和最高值分别出现在4月和1月, 年平均值为(57.02±4.00) mg · L-1;Fl-M处理最低值和最高值分别出现在1月和5月, 年平均值为(3634.02±240.86) mg · L-1;Sw-M处理最低值和最高值分别出现在1月和9月, 年平均值为(15575.84±1129.50) mg · L-1.多因素方差分析表明, 盐度、植物、时间均对河口湿地间隙水Cl-浓度有显著影响(表 3), 短叶茳芏湿地间隙水Cl-浓度显著大于光滩(p <0.05).波动盐度及恒定咸水处理均显著增加了湿地土壤间隙水Cl-浓度(p < 0.05), 短叶茳芏湿地中, 与恒定淡水处理相比, 波动盐度和恒定咸水处理下的间隙水Cl-浓度年均值分别增加了150.15和400.58倍;光滩湿地中, 相对于恒定淡水处理, 波动盐度和恒定咸水处理下的间隙水Cl-浓度年均值分别增加了62.73和272.16倍.
 |
| 图 6 恒定和波动盐度处理下间隙水Cl-、SO42-浓度的时间变化 Fig. 6 Temporal variations of porewater Cl-, SO42- concentration under constant and fluctuating salinity treatments |
间隙水SO42-浓度月动态见图 6.短叶茳芏湿地中, Fw-V处理最低值和最高值分别出现在6月和4月, 年平均值为(59.60±6.96) mg · L-1;Fl-V处理最低值和最高值分别出现在7月和5月, 年平均值为(333.05±46.30) mg · L-1;Sw-V处理最低值和最高值分别出现在1月和6月, 年平均值为(1534.31±94.09) mg · L-1.光滩中, Fw-M处理最低值和最高值分别出现在8月和3月, 年平均值为(50.13±4.14) mg · L-1;Fl-M处理最低值和最高值分别出现在7月和5月, 年平均值为(434.40±33.53) mg · L-1;Sw-M处理最低值和最高值分别出现在1月和7月, 年平均值为(2229.88±169.50) mg · L-1.多因素方差分析表明, 盐度、植物、时间均对河口湿地间隙水SO42-浓度有显著影响(表 3), 光滩间隙水SO42-浓度显著大于短叶茳芏湿地(p <0.05).波动盐度及恒定咸水处理均显著增加了湿地土壤间隙水SO42-浓度(p < 0.05), 短叶茳芏湿地中, 与恒定淡水处理相比, 波动盐度和恒定咸水处理下的间隙水SO42-浓度年均值增加了4.59和23.40倍;光滩湿地中, 相对于恒定淡水处理, 波动盐度和恒定咸水处理下的间隙水SO42-浓度年均值增加了7.67和43.48倍.
间隙水NH4+-N浓度月动态见图 7.短叶茳芏湿地中, Fw-V处理最低值和最高值分别出现在9月和1月, 年平均值为(1.06±0.36) mg · L-1;Fl-V处理最低值和最高值分别出现在9月和2月, 年平均值为(1.27±0.37) mg · L-1;Sw-V处理最低值和最高值分别出现在3月和9月, 年平均值为(7.02±1.04) mg · L-1.光滩中, Fw-M处理最低值和最高值分别出现在6月和7月, 年平均值为(4.39±0.41) mg · L-1;Fl-M处理最低值和最高值分别出现在5月和2月, 年平均值为(6.18±0.39) mg · L-1;Sw-M处理最低值和最高值分别出现在1月和8月, 年平均值为(17.17±0.89) mg · L-1.多因素方差分析表明, 盐度、植物、时间均对湿地间隙水NH4+-N浓度有显著影响(表 3), 光滩间隙水NH4+-N浓度显著大于短叶茳芏湿地(p <0.05).波动盐度及恒定咸水处理均显著增加了湿地土壤间隙水NH4+-N浓度(p < 0.05), 短叶茳芏湿地中, 与恒定淡水处理相比, 波动盐度和恒定咸水处理下的间隙水NH4+-N浓度年均值分别增加了0.19和5.62倍;光滩湿地中, 相对于恒定淡水处理, 波动盐度和恒定咸水处理下的间隙水NH4+-N浓度年均值分别增加了0.41和2.91倍.
 |
| 图 7 恒定和波动盐度处理下间隙水NH4+-N、NO3--N、TN浓度的时间变化 Fig. 7 Temporal variations of porewater NH4+-N, NO3--N, TN concentration under constant and fluctuating salinity treatments |
间隙水NO3--N浓度月动态见图 7.短叶茳芏湿地Fw-V、Fl-V和Sw-V处理下的间隙水NO3--N浓度最高值均出现在2月, Fw-V处理的最低值出现在9月, Fl-V和Sw-V处理的最低值均出现在8月, Fw-V、Fl-V和Sw-V处理的年均值分别为(1.11±0.41)、(1.15±0.42)、(1.10±0.42) mg · L-1;光滩中, Fw-M、Fl-M、Sw-M处理下的间隙水NO3--N浓度最高值均出现在2月, Fw-M和Fl-M处理的最低值均出现在9月, Sw-M处理的最低值出现在6月, Fw-M、Fl-M、Sw-M处理的年均值分别为(1.35±0.44)、(1.03±0.41)、(1.02±0.42) mg · L-1.不同处理下间隙水NO3--N浓度差异并不显著(p>0.05).
间隙水TN浓度月动态见图 7.短叶茳芏湿地Fw-V处理的最低值和最高值分别出现在7月和2月, 年平均值为(2.62±0.50) mg · L-1;Fl-V处理的最低值和最高值分别出现在6月和2月, 年平均值为(2.54±0.42) mg · L-1;Sw-V处理的最低值和最高值分别出现在3月和9月,年平均值为(8.85±1.04) mg · L-1.光滩中, Fw-M处理的最低值和最高值分别出现在6月和9月, 年平均值为(5.47±0.46) mg · L-1;Fl-M处理的最低值和最高值分别出现在5月和3月, 年平均值为(6.70±0.38) mg · L-1;Sw-M处理的最低值和最高值分别出现在1月和9月, 年平均值为(17.83±1.00) mg · L-1.多因素方差分析表明, 盐度、植物、时间均对河口湿地间隙水TN浓度有显著影响(表 3), 光滩间隙水TN浓度显著大于短叶茳芏湿地(p < 0.05), 此外, 恒定咸水处理显著大于波动盐度及恒定淡水处理(p < 0.05), 但波动盐度与恒定淡水处理之间差异不显著(p>0.05).
3.5 间隙水溶解性CH4浓度与间隙水理化因子、土壤温度的相关性间隙水溶解性CH4浓度与间隙水pH值、NH4+-N、NO3--N、TN、Cl-、SO42-浓度及气温的相关性见表 4.短叶茳芏湿地及光滩间隙水溶解性CH4浓度均与间隙水pH值呈显著正相关(p < 0.01), 与间隙水NH4+-N、NO3--N、TN浓度呈显著负相关(p < 0.05), 与间隙水Cl-、SO42-浓度同样呈显著负相关(p < 0.01), 此外, 光滩间隙水溶解性CH4浓度还与气温呈显著正相关性(p < 0.05).
| 表 4 间隙水溶解性CH4浓度与环境因子的Pearson相关性分析 Table 4 Pearson′s correlation coefficient between porewater dissolved CH4 concentration and environmental factors |
为进一步探讨各环境因子对间隙水溶解性CH4浓度的贡献大小, 在去除共线性后, 通过多元回归分析中的逐步回归法建立短叶茳芏湿地和光滩间隙水溶解性CH4浓度(分别为YV、YM)与间隙水pH值(X1)、NH4+-N(X2)、NO3--N(X3)、TN(X4)、Cl-(X5)、SO42-(X6)浓度、气温(X7)之间的最优回归方程(表 5).结果发现, 短叶茳芏湿地间隙水pH值对间隙水溶解性CH4浓度贡献更大, 光滩间隙水NH4+-N浓度及气温对间隙水溶解性CH4浓度贡献更大.
| 表 5 间隙水溶解性CH4浓度与环境因子的多元回归线性拟合方程 Table 5 Multiple regression equations between porewater dissolved CH4 concentration and environmental factors |
本研究结果表明, 持续恒定盐度增加处理和波动短期盐度增加处理均显著降低了河口淡水感潮湿地间隙水溶解性CH4浓度, 且在波动短期盐度处理下, 短叶茳芏湿地间隙水溶解性CH4浓度下降的幅度明显大于光滩.进一步分析发现, 导致以上结果的主要原因在于:恒定咸水和波动盐度增加处理下, 盐分输入一方面提高了间隙水Cl-、SO42-、NH4+-N、TN浓度, 另一方面降低了间隙水pH值, 继而对湿地间隙水溶解性CH4浓度产生抑制作用.
有研究发现, 长期水淹条件下形成的厌氧环境中, 沉积物间隙水中的无机氮主要以氨态形式出现(王圣瑞等, 2006; 陆健健, 2006);厌氧条件更利于反硝化作用的进行, 使得间隙水中NH4+-N浓度增加(王盼盼等, 2014), 此外, 沉积物有机质的矿化也会促使沉积物中NH4+-N释放(闫兴成等, 2018).本研究中, 间隙水NH4+-N浓度与间隙水溶解性CH4浓度之间存在显著的负相关关系(表 4).以往研究发现, 外源氮输入会抑制河口咸草湿地土壤CH4产生潜力(王维奇等, 2010), 无机氮输入(NH4+-N)会抑制三江平原小叶章湿地土壤CH4的产生(陶宝先等, 2016), 土壤细菌活性会因NH4+-N的添加而降低(Soderberg et al., 2004), 进而抑制土壤CH4的产生.
本研究中, 恒定咸水及波动盐度增加处理带来大量的Cl-和SO42-, 水体通过下渗作用促使间隙水Cl-和SO42-浓度增加.相关分析表明(表 4), 间隙水Cl-和SO42-浓度与间隙水溶解性CH4浓度之间存在显著的负相关性, 这与大多数研究结果一致.较高的SO42-浓度通过还原作用降解有机碳产生的能量, 并通过与产甲烷菌竞争醋酸、氢等底物及产生毒害和改变环境因子等方式对CH4代谢过程产生抑制作用(王进欣等, 2011; Poffenbarger et al., 2011; 王维奇等, 2012; Marton et al., 2012); 并且厌氧条件下, SO42-会通过促进厌氧甲烷氧化的过程, 抑制甲烷排放(Knittel et al., 2009).而较高的Cl-浓度会影响植物及微生物生长, 具有一定的胁迫作用(张子川等, 2015; 王纯, 2015), 主要通过离子毒害、渗透胁迫、营养失衡及盐胁迫的次级反应等途径影响土壤产甲烷菌所需的底物供应及其活性(Baldwin et al., 2006; 杨平等, 2015), 进而抑制湿地间隙水溶解性CH4浓度.
与此同时, 盐分输入带来的Cl-和SO42-等强酸性离子可使pH值降低(侯晓娟等, 2010; 冯小平等, 2015).以往研究表明, 产甲烷菌可承载的土壤pH为5.5~9.0, 最适范围在6.8~7.2, 超出最适范围, CH4排放较低(牟晓杰等, 2012).而本研究中, 恒定咸水及波动盐度的处理均明显降低了湿地间隙水pH值, 并且不同处理下的短叶茳芏湿地间隙水pH值普遍低于光滩, 特别是在恒定咸水处理下, 其间隙水pH值低于产甲烷菌的可承载范围.
尽管恒定咸水和波动盐度增加处理可引起间隙水NH4+-N、TN、Cl-、SO42-浓度的升高及间隙水pH值的降低, 会降低间隙水溶解性CH4浓度, 但通过多元回归分析发现, 短叶茳芏湿地间隙水溶解性CH4浓度更多受间隙水pH值影响, 而光滩间隙水溶解性CH4浓度则受间隙水NH4+-N浓度影响更大.
此外, 值得注意的是, 温度对湿地甲烷的产生、氧化和传输均有重要影响, 土壤产甲烷能力会随温度升高而增加(Rooneyvarga et al., 2007; Eriksson et al., 2010; 沙晨燕等, 2015), 温度较低时, 植物和微生物活性被削弱, 甲烷产生量较低(李玲等, 2012).本研究中, 不论是短叶茳芏湿地抑或是光滩, 其间隙水溶解性CH4浓度均随温度的升高而增加, 特别进入夏季以后, 但温度对于短叶茳芏湿地间隙水溶解性CH4浓度的影响并不显著, 对于光滩间隙水溶解性CH4浓度的影响更为显著.有研究表明, 土壤或沉积物厌氧区产生的CH4, 可以通过植被根系对水分的吸收或者扩散进入植物体, 被植物运输并释放到大气当中(Nisbet et al., 2009; Rice et al., 2010; Askaer et al., 2011).而短叶茳芏湿地中植物的存在, 虽然一方面为间隙水甲烷的产生提供了底物, 提高了有植被覆盖湿地间隙水溶解性CH4浓度, 但更多的表现为促进间隙水溶解性CH4的排放, 特别是4月以后, 短叶茳芏进入生长旺季, 加之其他环境因子的影响, 因此削弱了温度对于短叶茳芏湿地间隙水溶解性CH4浓度的影响.
综合以上影响因素, 导致恒定咸水及波动盐度处理均对河口湿地间隙水溶解性CH4浓度产生了明显的抑制作用.
5 结论(Conclusions)1) 恒定咸水及波动盐度增加处理均显著降低了河口淡水感潮湿地间隙水溶解性CH4浓度, 并且恒定咸水处理的抑制作用更强, 而波动盐度增加对于短叶茳芏湿地间隙水溶解性CH4浓度的抑制效果明显高于光滩.
2) 短叶茳芏湿地间隙水溶解性CH4浓度受间隙水pH值影响显著, 而光滩湿地间隙水溶解性CH4浓度则受间隙水NH4+-N浓度及气温影响显著.
3) 未来盐水入侵情景下, 河口淡水感潮湿地间隙水溶解性CH4浓度将下降, 且在盐度短期波动增加情景下, 河口淡水感潮沼泽湿地间隙水溶解性CH4浓度下降幅度显著大于光滩湿地.
Askaer L, Bo E, Friborg T, et al. 2011. Plant-mediated CH4, transport and C gas dynamics quantified in-situ in a Phalaris arundinacea-dominant wetland[J]. Plant and Soil, 343(1/2): 287–301.
|
Baldwin D S, Rees G N, Mitchell A M, et al. 2006. The short-term effects of salinization on anaerobic nutrient cycling and microbial community structure in sediment from a fresh water wetland[J]. Wetlands the Journal of the Society of the Wetland Scientists, 26(2): 455–464.
|
Chambers L G, Reddy K R, Osborne T Z. 2011. Short-term response of carbon cycling to salinity pulses in a freshwater wetland[J]. Soil Science Society of America Journal, 75(5): 2000–2007.
DOI:10.2136/sssaj2011.0026
|
Ding W X, Zhang Y H, Cai Z C. 2010. Impact of permanent inundation on methane emissions from a Spartina alterniflora coastal salt marsh[J]. Atmospheric Environment, 44(32): 3894–3900.
DOI:10.1016/j.atmosenv.2010.07.025
|
Ding W, Cai Z, Tsuruta H. 2005. Factors affecting seasonal variation of methane concentration in water in a freshwater marsh vegetated with Carex lasiocarpa[J]. Biology and Fertility of Soils, 41(1): 1–8.
DOI:10.1007/s00374-004-0812-9
|
Eriksson T, Ouml Quist M G, Nilsson M B. 2010. Production and oxidation of methane in a boreal mire after a decade of increased temperature and nitrogen and sulfur deposition[J]. Global Change Biology, 16(7): 2130–2144.
DOI:10.1111/gcb.2010.16.issue-7
|
冯小平, 王义东, 王博祺, 等. 2015. 盐分对湿地甲烷排放影响的研究进展[J]. 生态学杂志, 2015, 34(1): 237–246.
|
Geurts J J, Smolders A J, Banach A M, et al. 2010. The interaction between decomposition, net N and P mineralization and their mobilization to the surface water in fens[J]. Water Research, 44(11): 3487.
DOI:10.1016/j.watres.2010.03.030
|
侯晓娟, 徐明岗, 李冬初, 等. 2010. 长期施用含硫含氯化肥稻田土壤化学性质的演变特征[J]. 中国农业科学, 2010, 43(12): 2460–2468.
DOI:10.3864/j.issn.0578-1752.2010.12.009 |
IP CC. 2013. The Physical Science Basis.Working Group I Contribution to the Fifth Assessment Report of the Inter-governmental Panel on Climate Change[M]. Cambridge: Cambridge University Press.
|
姜欢欢, 孙志高, 王玲玲, 等. 2012. 秋季黄河口滨岸潮滩湿地系统CH4通量特征及影响因素研究[J]. 环境科学, 2012, 33(2): 565–573.
|
Kazunori M, Seiichi N, Takuji S, et al. 2010. Annual emissions of dissolved CO2, CH4, and N2O in the subsurface drainage from three cropping systems[J]. Global Change Biology, 16(2): 796–809.
|
Kiehn W M, Mendelssohn I A, White J R. 2013. Biogeochemical recovery of oligohaline wetland soils experiencing a salinity pulse[J]. Soil Science Society of America Journal, 77(6): 2205–2215.
DOI:10.2136/sssaj2013.05.0202
|
Knittel K, Boetius A. 2009. Anaerobic oxidation of methane:Progress with an unknown process[J]. Annual Review of Microbiology, 63(63): 311.
|
Konnerup D, Villamil C, Parra J P. 2014. Nitrous oxide and methane emissions from the restored mangrove ecosystem of the Ciénaga Grande de Santa Marta, Colombia[J]. Estuarine Coastal and Shelf Science, 140(Complete): 43–51.
|
Krauss K W, Whitbeck J L. 2012. Soil greenhouse gas fluxes during wetland forest retreat along the lower Savannah River, Georgia (USA)[J]. Wetlands, 32(1): 73–81.
DOI:10.1007/s13157-011-0246-8
|
Loáiciga H A, Pingel T J, Garcia E S. 2012. Sea water intrusion by sea-level rise:scenarios for the 21st century[J]. Groundwater, 50(1): 37–47.
DOI:10.1111/gwat.2012.50.issue-1
|
李玲, 仇少君, 刘京涛, 等. 2012. 土壤溶解性有机碳在陆地生态系统碳循环中的作用[J]. 应用生态学报, 2012, 23(5): 1407–1414.
|
刘剑秋, 曾从盛. 2010. 福建湿地及其生物多样性[M]. 北京: 科学出版社.
|
罗敏, 黄佳芳, 刘育秀, 等.一种滨海湿地滨海潮滩物孔隙水采样器湿地沉积物孔隙水采样器[P].中国.201621472027.1.2016-12-30
|
Marton J M, Herbert E R, Craft C B. 2012. Effects of salinity on denitrification and greenhouse gas production from laboratory-incubated tidal forest soils[J]. Wetlands, 32(2): 347–357.
DOI:10.1007/s13157-012-0270-3
|
牟晓杰, 刘兴土, 仝川, 等. 2012. 闽江河口短叶茳芏湿地CH4和N2O排放对氮输入的短期响应[J]. 环境科学, 2012, 33(7): 2482–2489.
|
Neubauer S C. 2013. Ecosystem responses of a tidal freshwater marsh experiencing saltwater intrusion and altered hydrology[J]. Estuaries and Coasts, 36(3): 491–507.
DOI:10.1007/s12237-011-9455-x
|
Nisbet R E R, Fisher R, Nimmo R H, et al. 2009. Emission of methane from plants[J]. Proceedings of the Royal Society B Biological Sciences, 276(1660): 1347.
DOI:10.1098/rspb.2008.1731
|
Pivni-ková, Rejmánková B, Snyder E, et al. 2010. Heterotrophic microbial activities and nutritional status of microbial communities in tropical marsh sediments of different salinities:the effects of phosphorus addition and plant species[J]. Plant and Soil, 336(336): 49–63.
|
Poffenbarger H J, Needelman B A, Megonigal J P. 2011. Salinity influence on methane emissions from tidal marshes[J]. Wetlands, 31(5): 831–842.
DOI:10.1007/s13157-011-0197-0
|
Rice A L, Butenhoff C L, Shearer M J, et al. 2010. Emissions of anaerobically produced methane by trees[J]. Geophysical Research Letters, 37(3): 202–217.
|
Rooneyvarga J N, Giewat M W, Duddleston K N, et al. 2007. Links between archaeal community structure, vegetation type and methanogenic pathway in Alaskan peatlands[J]. Fems Microbiology Ecology, 60(2): 240–251.
DOI:10.1111/fem.2007.60.issue-2
|
Roos L, Erika V D, Leonpm L, et al. 2007. How soil characteristics and water quality influence the biogeochemical response to flooding in riverine wetlands[J]. Biogeochemistry, 85(3): 289–302.
DOI:10.1007/s10533-007-9135-x
|
Soderberg K H, Baath E. 2004. The influence of nitrogen fertilization on bacterial activity in the rhizosphere of barley[J]. Soil Biology and Biochemistry, 36(1): 195–198.
DOI:10.1016/j.soilbio.2003.07.001
|
Sun Z, Jiang H, Wang L, et al. 2013. Seasonal and spatial variations of methane emissions from coastal marshes in the northern Yellow River estuary, China[J]. Plant and Soil, 369(1): 317–333.
|
沙晨燕, 谭娟, 王卿, 等. 2015. 不同类型河滨湿地甲烷和二氧化碳排放初步研究[J]. 生态环境学报, 2015, 24(7): 1182–1190.
|
陶宝先, 陈永金. 2016. 不同形态氮输入对湿地生态系统碳循环影响的研究进展[J]. 生态环境学报, 2016, 25(1): 162–167.
|
王纯.2015.模拟盐水入侵对闽江口淡水湿地温室气体产生与排放的影响[D].福州: 福建师范大学
http://cdmd.cnki.com.cn/Article/CDMD-10394-1015717779.htm |
王进欣, 王今殊, 钦佩, 等. 2011. 生源气体排放的潮周期动态研究:关键科学问题与不确定性[J]. 海洋湖沼通报, 2011(4): 134–143.
DOI:10.3969/j.issn.1003-6482.2011.04.019 |
王盼盼, 李玲玲, 陈洪涛, 等. 2014. 黄河口湿地沉积物间隙水中营养盐研究[J]. 海洋湖沼通报, 2014(4): 97–103.
|
王维奇, 曾从盛, 仝川, 等. 2012. 闽江河口潮汐湿地二氧化碳和甲烷排放化学计量比[J]. 生态学报, 2012, 32(14): 4396–4402.
|
王维奇, 曾从盛, 仝川. 2010. 闽江河口湿地土壤甲烷产生潜力动态及对氮输入的响应[J]. 农业系统科学与综合研究, 2010, 26(2): 209–213.
DOI:10.3969/j.issn.1001-0068.2010.02.016 |
Weston N B, Dixon R E, Joye S B. 2006. Ramifications of increased salinity in tidal freshwater sediments:Geochemistry and microbial pathways of organic matter mineralization[J]. Journal of Geophysical Research Atmospheres, 111(G1): 689–699.
|
闫兴成, 王明玥, 许晓光, 等. 2018. 富营养化湖泊沉积物有机质矿化过程中碳、氮、磷的迁移特征[J]. 湖泊科学, 2018, 30(2): 306–313.
|
杨平, 张子川, 杜威宁, 等. 2015. 河口盐度梯度下短叶茳芏沼泽湿地土壤间隙水溶解性甲烷时空特征[J]. 环境科学, 2015, 36(10): 3633–3640.
|
曾从盛, 王维奇, 仝川. 2008. 不同电子受体及盐分输入对河口湿地土壤甲烷产生潜力的影响[J]. 地理研究, 2008, 27(6): 1321–1330.
DOI:10.3321/j.issn:1000-0585.2008.06.011 |
张子川, 杨平, 仝川. 2015. 盐分对河口淡水、微咸水沼泽湿地土壤甲烷产生潜力的影响[J]. 生态学报, 2015, 35(24): 8075–8084.
|