 2019, Vol. 39
2019, Vol. 39



硫(S)作为继氮、磷、钾之后的第4位营养元素在植物生长发育及代谢过程中不可或缺, 如参与蛋白质及氨基酸合成、控制光合作用中碳水化合物代谢、影响呼吸作用和抗逆性等(陆景陵, 2003).缺硫可导致植物代谢紊乱、生长发育受阻乃至死亡, 其对维持生态系统的健康发展至关重要(李新华等, 2007a).滨海湿地是陆地和海洋之间过渡的重要生态类型, 海陆作用强烈, 是海岸带地区响应全球变化和人类活动较为敏感的重要生态系统之一(Simas et al., 2001), 同时也是极易受到外来生物入侵的重要生态系统(Mccary et al., 2016).
互花米草(Spartina alterniflora)作为我国沿海地区淤泥质滩涂的主要入侵植物(刘春悦等, 2009), 具有耐盐性、耐淹性、根系发达、繁殖力强等特点(Seliskar et al., 2002), 该植物抗氧化系统在抵御硫胁迫过程中比芦苇、碱蓬等植物响应能力强(胡楚琦等, 2015), 对高硫环境具有很强的适应性, 因此, 研究硫含量分布特征可为揭示互花米草扩张机制提供理论基础.当前, 国内尽管对湿地硫营养元素进行了不少研究, 包括土壤硫形态分布(Zhou et al., 2007)、含硫气体释放特征(李新华等, 2014; 2015)、硫生物循环状况评估(Sun et al., 2013)等, 但有关湿地互花米草入侵下硫含量的研究仍非常有限, 主要集中在江苏盐城(Zhou et al., 2009)、闽江河口(何涛等, 2016)等区域, 研究中重点关注了土壤硫含量的空间分布特征.由于互花米草具有高生物量及高生产力特征, 随着互花米草生长时期变化会对硫含量产生影响, 而当前从生长阶段开展对湿地土壤、互花米草硫含量的研究还鲜有报道.
胶州湾湿地是山东半岛最大的河口海湾型湿地, 已被列入《国家重要湿地名录》, 对维护区域生态安全发挥着重要作用.该区域湿地总面积约17.76×104 hm2(訾园园等, 2016).洋河入湾口湿地是典型的河口湿地, 互花米草于2009年已有零星分布, 目前洋河口南岸湿地互花米草长势旺盛, 面积约为77.2 hm2, 并持续向低潮滩蔓延.目前的研究表明, 互花米草入侵不仅对湿地生态系统的过程、功能与服务有显著影响(Ehrenfeld, 2003), 也可能对生物地球化学过程中生源要素存在潜在影响(Seliskar et al., 2004).当前关于互花米草入侵胶州湾湿地的研究主要集中在土壤碳、氮、磷等大量元素方面(訾园园等, 2016; 谢文霞等, 2014; 苗萍等, 2017), 而对硫或其他中微量元素的关注还十分不足.因此, 本研究选取胶州湾洋河入湾口互花米草潮滩及光滩为研究对象, 对比分析2个样区土壤总硫(Total sulfur, TS)、有效硫(Available sulfur, AS)的时空分布特征及其影响因素, 研究互花米草在不同生长阶段的TS累积分配特征与土壤硫含量的关系, 旨在进一步了解互花米草入侵后滨海湿地营养元素硫的生物地球化学循环, 为胶州湾湿地的可持续发展提供科学依据.
2 材料与方法(Materials and methods) 2.1 研究区概况研究区位于山东半岛胶州湾(36°05′~36°15′N, 120°03′~120°25′E)洋河入湾口(图 1).该区域属于暖温带季风气候, 年平均气温为12.2 ℃, 8月平均气温为25.5 ℃, 1月平均气温为-1.2 ℃, 无霜期在220 d左右, 年平均降雨量为775.6 mm, 降水量年内分布不均, 且主要集中在夏季.胶州湾沿岸潮汐为典型的半日潮, 平均潮差2.71 m, 最大潮差6.87 m.区内土壤pH为7.01~8.28, 土壤类型主要是粘质土和沙质土.据实地考察, 研究区互花米草群落初春开始返青, 9月以前总体处于营养生长期;9月中旬到10月中旬处于开花期;11月中旬后进入非生长季, 植株开始泛黄, 逐渐枯萎.
 |
| 图 1 研究区及采样点位置 Fig. 1 Study area and sampling sites |
依据典型性、代表性、一致性和可行性原则, 于2016年6月(营养生长期)、10月(生殖生长期)、12月(非生长期), 基于遥感影像解译和实地调查数据, 采用时空互代法选取洋河入湾口湿地水文条件基本一致、土壤与成土母质类型相同的互花米草区(H区, 代表入侵后, 采样点入侵年限为5年)和无植被光滩区(M区, 代表入侵前)进行采样(图 1).垂直于岸线设置3条样线, 每条样线均能被大小潮淹没, 分别在H和M区利用土钻采集3个0~60 cm深土壤柱状样品, 作为重复, 共分为6层.取土后立即将土壤装入密封袋中带回实验室.土壤样品在自然条件下风干, 除去肉眼可见的植物枯落物、碎石等杂质, 研磨过100目(0.149 mm)筛后, 储存于自封袋中待测.在互花米草样区, 采集土壤样品的同时, 随机选取3个50 cm×50 cm样方, 用刀沿地面割下植物地上部分, 地下部分采用挖掘法, 将样方内所有地下根系全部挖出, 收集后带回实验室.植物清洗干净后分离为根、茎、叶, 放入烘箱105 ℃杀青, 80 ℃烘干至恒重, 磨碎、过100目筛后, 装入自封袋中待测.3个月土壤、植物样品分别为108个和27个.测得供试土壤的基本理化性质见表 1.
| 表 1 供试土壤理化性质(0~60 cm) Table 1 Physicochemical properties of soil tested (0~60 cm) |
| 表 3 土壤TS垂直分布变异性 Table 3 Vertical variability of TS contents in soils |
| 表 5 土壤AS垂直分布变异性 Table 5 Vertical variability of AS contents in soils |
土壤和植物总硫(TS)的测定采用酸消解-BaSO4比浊法(佩奇等, 1991);土壤有效硫(AS)采用CaCl2浸提-BaSO4比浊法测定(郝庆菊等, 2003);总铁采用酸消解-邻菲罗啉光度法测定(刘光崧, 1996);土壤有机质、EC、pH、含水量、容重分别采用K2CrO7-H2SO4外加热法、电导法(水土质量比5:1)、电位法(水土质量比5:1)、烘干法、环刀法测定(鲍士旦, 2000).
2.3 指标计算植物硫累积系数(AFS)按以下公式计算(李新华, 2007):
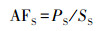
|
式中, PS表示植物器官中TS含量(g·kg-1), SS表示土壤TS含量(g·kg-1).
2.4 数据处理采用Excel 2007和Origin 9进行数据处理与图表制作.运用SPSS 20.0进行土壤TS、AS与环境因子间相关性分析和逐步线性回归分析, 并用单因素方差分析法(One-way ANOVA)及多因素方差分析法(Multi-way ANOVA)进行数据差异性显著检验(α=0.05).
3 结果与分析(Results and analysis) 3.1 土壤硫含量 3.1.1 土壤TS含量采样区土壤TS含量变化范围较大, 为0.44~2.87 g·kg-1, 互花米草入侵后及光滩0~60 cm土壤TS平均含量分别为(1.53±0.24) g·kg-1、(1.13±0.09) g·kg-1, 入侵后TS含量增幅为35.40%, 增加显著(n=54, p < 0.05).同一时期, 土壤(0~60 cm)TS平均含量均呈现为入侵后大于光滩(表 2).入侵后各月土壤(0~60 cm)TS平均含量表现为10月>12月>6月, 且10月显著高于6月、12月(n=18, p < 0.05);光滩表现为6月>10月>12月, 但3个月份差异不明显(n=18, p>0.05).在10月, 互花米草潮滩土壤TS含量显著高于光滩(n=18, p < 0.05), 6月、12月不同湿地间TS含量差异不显著(n=18, p>0.05, 表 3).
| 表 2 土壤TS水平分布变异性 Table 2 Horizontal variability of TS contents in soils |
| 表 4 土壤AS水平分布变异性 Table 4 Horizontal variability of AS contents in soils |
2种湿地类型土壤TS含量在水平和垂直方向上均存在一定分异(图 2).Cambardella等(1994)对变异系数(CV)的分级标准为:CV < 10%为弱变异性, 10%~100%为中等变异性, CV>100%为强变异性.水平方向上, 2种湿地类型0~30 cm及50~60 cm相同土层间TS含量差异不显著(n=9, p>0.05), 但30~50 cm土层间差异显著(n=9, p < 0.05).互花米草入侵后0~10 cm表层土壤TS含量与30~60 cm各土层TS含量存在显著性差异(n=9, p < 0.05), 光滩各土层差异不显著(n=9, p>0.05, 表 2).
就不同时期而言, 互花米草入侵后明显改变了湿地土壤TS含量的分布特征和变异程度(图 2、表 3).6月, 互花米草入侵后湿地土壤TS含量在垂直方向上总体呈先增后减的趋势, 并于40~50 cm土层取得最大值;而互花米草入侵前的光滩湿地土壤TS含量的垂直变化特征与之相反.10月互花米草湿地各层土壤TS含量均高于光滩湿地, 分别在40~50 cm和10~20 cm土层取得最大值, 且在垂直方向上的变化分别属于中等变异性和弱变异性.与之相比, 12月两种类型湿地0~10 cm土层TS含量明显低于其他月份, 且入侵前的光滩湿地土壤TS含量略高于互花米草湿地, 而二者在20~60 cm各层的TS含量总体变化较小, 且互花米草入侵后湿地土壤TS含量明显高于光滩湿地.整体而言, 互花米草入侵后各土层TS含量及垂直变异程度大于入侵前.
 |
| 图 2 土壤TS含量垂直分布特征 Fig. 2 Vertical distribution of TS contents in soils |
土壤AS是可供植物直接吸收利用的部分, 其含量变化与植物生长密切相关.互花米草潮滩及光滩土壤(0~60 cm)AS平均含量分别为(0.75±0.04) g·kg-1和(0.53±0.02) g·kg-1, 各占土壤TS含量的49.02%和46.90%.入侵后不同时期土壤AS含量均显著高于光滩(n=18, p < 0.05).2种湿地类型土壤AS平均含量均为10月最高, 12月次之, 6月最低, 且垂直变异程度整体较弱(表 5).从水平方向上, 不同湿地各土层AS含量均属中等变异性, 除10~20 cm土层, 不同湿地相同土层间AS含量均差异显著(n=9, p < 0.05).同一湿地, 各土层间AS含量差异不显著(n=9, p>0.05, 表 4).
尽管不同类型湿地同一时期土壤AS含量垂直分布特征较为相似, 但互花米草入侵后湿地土壤AS含量整体高于光滩湿地(图 3).6月互花米草潮滩和光滩土壤AS含量均随深度增加而降低, 且0~30 cm及50~60 cm各土层AS含量均差异显著(n=3, p < 0.05), 入侵后垂直变异性属于中等变异性, 光滩则属于弱变异.10月, 2种湿地类型AS含量垂直变异性均较弱, 均在0~10 cm土层出现最大值(分别为0.98、0.71g·kg-1), 但互花米草潮滩在10~20 cm土层出现最低值(0.82 g·kg-1), 40~50 cm土层AS含量略有增加.12月, 2种湿地类型AS含量均在0~10 cm土层出现最低值, 在10~20 cm土层出现最大值(分别为0.86、0.71 g·kg-1), 20~60 cm各土层AS含量之间差异不显著(n=3, p>0.05), 光滩的垂直变异性大于互花米草潮滩.
 |
| 图 3 土壤AS含量垂直分布特征 Fig. 3 Vertical distribution of AS contents in soils |
入侵种互花米草根、茎、叶TS含量均值最高的月份均为6月(3.15 g·kg-1), 与12月呈差异显著(n=9, p < 0.05), 但6月与10月、10月与12月之间TS含量差异均不显著(n=9, p>0.05)(图 4).互花米草各器官TS平均含量在6月、10月均表现为叶>根>茎, 且同一月份叶TS含量均显著高于茎(n=3, p < 0.05);12月表现为根>叶>茎, 植物根与叶TS含量间无显著性差异(n=3, p>0.05), 根TS含量显著高于茎(n=3, p < 0.05).植物同种器官内, 6、10、12月的茎、叶TS含量随时间推移而降低, 6月根TS含量显著高于10月(n=3, p < 0.05), 呈现为6月>12月>10月, 可见根TS含量波动最大(图 4).
 |
| 图 4 互花米草各器官TS含量(不同大写字母表示同一器官不同月份间差异显著, 不同小写字母同一月份不同器官间差异显著(p < 0.05)) Fig. 4 Average TS content in different organs of S. alterniflora |
整体而言, 互花米草群落硫累积量最高值出现在10月, 其次为12月, 6月最低(表 6).同一时期, 茎为硫累积量最高的器官.同一器官内, 互花米草群落根、茎、叶的AFs值均呈现为6月>12月>10月.同一时期, 6月、10月均表现为叶的AFs值最高, 根次之, 茎最低, 12月呈现为根最高, 叶次之, 茎最低(表 6).
| 表 6 互花米草不同器官硫累积量与AFs值 Table 6 Sulfur accumulations and AFs values in different organs of S. alterniflora |
土壤中TS含量变化较大, 一般在0.03~10.00 g·kg-1之间, 其平均值约为0.70 g·kg-1(Lindsay, 1979).研究区土壤(0~60 cm)TS均值为1.33 g·kg-1, 约为世界土壤含S量平均值的2倍.对比发现, 同一研究区域互花米草湿地土壤TS含量明显高于光滩和其他植被类型湿地(表 7).原因可能在于互花米草可依靠生理学过程储存高浓度的硫, 使得其生长环境中的硫浓度保持在较高状态.对比研究表明, 本研究区湿地土壤TS含量总体处于较低水平.已有研究表明, 土壤有机质是影响土壤硫含量的重要因素之一(Lamers et al., 2013).而研究区位于北温带季风气候区, 年平均气温低, 地表有机物累积能力、有机物的分解速度、残体中硫归还速率均较低, 从而使得湿地土壤TS含量总体较低.另外, 研究区互花米草潮滩TS含量低于闽江河口, 不同生境条件下互花米草对环境中硫吸收与累积能力的差异也会影响湿地土壤硫含量.目前, 关于滨海湿地土壤AS的研究相对较少, 但本研究湿地土壤AS含量明显高于闽江河口, 原因可能为研究区每日受大小潮淹没, 海水补给湿地大量硫酸盐, 且土壤呈碱性(表 1), 水溶性硫具有较高水平, 而水溶性硫是最易被植物吸收利用的硫形态.与我国沿海地区互花米草湿地相比, 国外报道的互花米草湿地土壤TS含量普遍处于较高水平, 原因可能有两方面:一方面可能与土壤硫库储量有关, 已有研究表明, 不同区域沉积环境和海洋SO42-赋存量存在较大差异(Ferreira et al., 2007), 国外相关研究区域土壤和海水环境中硫赋存量较多, 使得其湿地土壤TS高于我国滨海湿地.另一方面, 互花米草具有高遗传分化和基因渗入能力, 生境差异可造成互花米草50%的生态型差异(Perkins et al., 2002; Richards et al., 2005), 而在我国, 不同空间尺度上互花米草种群遗传多样性都相对较低(邓自发等, 2006).
| 表 7 不同研究区域互花米草湿地硫含量对比 Table 7 Comparison of sulfur contents in S. alterniflora wetland in different study areas |
采用多因素方差分析(Multi-way ANOVA)对研究区植被类型、植物生长月份、土层深度等因素作为固定因子进行检验可知(表 8), 植被类型、植被类型与月份的交互作用对土壤TS均存在显著影响(p < 0.05), 土壤TS的变异被3种固定因子及它们的交互作用解释的部分占37.90%(R2=0.379);植被类型及月份对土壤AS均存在显著的单独影响(p < 0.05), 固定因子间交互作用对土壤AS影响不显著(p>0.05), 土壤AS的变异被3种控制因子及它们的交互作用解释的部分占60.30%(R2=0.603).总体而言, 湿地土壤硫含量受植被类型及月份的影响较大.
| 表 8 主体间效应的检验 Table 8 Tests of between-subjects effects |
群落演替尤其是植物入侵可能会显著影响土壤理化性质(Chacón et al., 2009).一方面植被通过自身吸收、累积与返还影响湿地土壤硫含量;另一方面, 互花米草入侵后可导致湿地土壤理化性质改变而间接影响.本研究主要对互花米草入侵后湿地土壤TS、AS含量与非生物因素及植物吸收累积能力2个方面进行相关性分析及逐步回归分析(表 9、表 10).本研究表明, 有机质是影响互花米草潮滩土壤TS、AS分布的重要因子(表 9), 原因可能为互花米草残体返还土壤致使有机质含量增加, 有机质中含有一定数量的S官能团(林慧娜等, 2009)促使硫累积, 而有机质的含量及分解能力影响着土壤AS含量的高低.光滩土壤硫与有机质无明显相关性(p>0.05), 原因可能是光滩有机质含量变化不大, 对土壤硫影响较小.互花米草入侵后土壤EC也是影响土壤AS的关键因素(表 10).互花米草是泌盐植物, 通过吸收、累积盐分而使土壤盐分减少, 但采样区盐分主要来自潮水的输入, 盐分通过植物泌盐腺返还加之植物对潮水的滞留, 因而使得入侵区可溶性盐显著大于光滩(n=54, p < 0.05).可溶性盐通过影响微生物活动来影响凋落物分解速率和硫的矿化作用, 从而影响硫的含量(Lu et al., 2016a), 其与土壤硫呈显著正相关(p < 0.05), 这与曾从盛等(2010)研究结果相一致.土壤含水量会影响有机质合成与分解、硫元素迁移转化及植被生长(许秀丽等, 2014).影响光滩土壤TS、AS含量的关键因素为含水量(表 10), 这可能与研究区水动力剧烈, 冲淤作用较为频繁, 以及潮水的周期性循环有关.互花米草入侵后, 土壤硫与含水量、容重均无明显关系(p>0.05), 这可能与互花米草群落环境因子更加复杂及其消浪缓流功能有关.
| 表 9 硫素与环境因子的相关系数 Table 9 Correlation coefficients of sulfur contents and environmental factors |
| 表 10 土壤硫素与环境因子的逐步回归分析 Table 10 Stepwise regression analyses of the relationships between soil sulfur and environmental factors |
由表 9可知,研究区土壤TS均与总铁呈显著正相关(p < 0.01), 原因可能是土壤中S与Fe元素结合生成FeS、FeS2等硫化物(于君宝等, 2014), 从而影响土壤TS的分布.2种类型湿地土壤pH与土壤AS呈显著负相关关系(p < 0.01), 这是因为河口湿地硫的主要赋存形态硫酸盐受pH的影响较大(单孝全等, 1991).
互花米草TS含量、AFs与土壤AS之间存在显著负相关性(p < 0.05), 植物生物量、S累积量与土壤AS呈显著正相关(p < 0.05), 可见植物因素与土壤AS含量分布密切相关(表 9).
4.3 互花米草入侵对湿地土壤TS、AS时空分布的影响互花米草在不同生长期对土壤硫存在一定影响, 这就使得开展不同生长阶段互花米草对土壤硫含量动态变化的影响研究显得尤为必要.研究区受植被类型影响显著, 有植被覆盖湿地土壤硫含量明显大于无植被光滩, 这可能与植被对环境中硫的吸收与累积能力有关.相关研究表明, 互花米草是C4植物, C4植物的CO2补偿点较低, 提高了强光、高温下的光合速率, 具有较高的生产力(牛书丽等, 2004).相比其他湿地草本植物而言, 互花米草对氮养分和光等有较高的利用效率, 其叶面积指数更大, 生长期更长, 净光合速率和净初级生产力更高(Jiang et al., 2009).湿地植物对营养元素的累积特征随生长节律表现出明显的季节动态(郭雪莲等, 2007).本研究表明, 互花米草入侵后土壤TS及AS在6月的含量相比于10月、12月最低, 10月含量最高(表 3、表 5), 植物TS含量则是6月、10月、12月逐步降低(图 4), 且互花米草入侵后各月份湿地土壤TS和AS含量均高于入侵前的光滩湿地.原因可能在于6月是互花米草的生长旺季, 对养分的累积能力较强, 其需要足够的硫养分来维持其较高的光合效率(胡楚琦等, 2015);而本研究中6月互花米草根、茎、叶的AFs值相较其他月份最高(表 6), 且AFs与土壤AS呈显著负相关(表 9), 因此, 入侵后土壤AS和TS含量较其他月份较低.而互花米草入侵后较强的养分累积能力和生产力导致其可以从环境中固持大量的硫养分(Zhou et al., 2009), 加之其可通过根系吸收土壤中可溶性S2-而将其氧化成稳定的SO42-来合成自身组成部分, 从而导致植物硫库储量增加(夏露, 2015; Carlson et al., 1982).互花米草进入成熟期(10月)后, 根、茎、叶的AFs值相较其他月份最低, 植物对硫的累积能力相对较低, 而湿地枯落物分解也是湿地生态系统的重要功能之一, 通过枯落物分解, 营养元素归还土壤, 不断被植物循环利用, 维持着生态系统的平衡(郭雪莲等, 2007).研究区上一年植物立枯物基本倒伏, 部分植物残体的返还, 加之互花米草根系发达, 大量死根分解可将硫释放到不同土层, 从而导致入侵后土壤硫化物浓度及硫库储量增加.12月互花米草潮滩土壤TS和AS含量均高于光滩, 但互花米草入侵后12月土壤TS、AS含量较10月略有下降.一方面是因为研究区冬季气温较低, 土壤表层甚至出现结冰现象, 微生物活性弱, 植物残体和有机质分解缓慢, 土壤硫补充不及时;另一方面植物根TS含量显著高于10月, 互花米草是多年生植物, 在12月, 根的AFs值最高, 根器官累积大量硫以供互花米草冬季消耗(李新华等, 2007b).
从垂直分布上看, 2种湿地类型表层土壤TS含量较少, 这与Lu等(2016b)的研究结果一致, 表层土壤硫易随径流流失或产生淋溶作用.互花米草入侵后, 10月土壤TS含量达到最大值, 入侵后有机质是影响土壤硫含量的关键因子(表 10), 互花米草生长年限较长, 根系集中于0~30 cm土层, 甚至能延伸到50~100 cm土层, 致使土壤深层有机质含量增多, 从而使40~50 cm土层TS含量有所累积.光滩0~10 cm土壤TS含量出现最大值, 而含水量为影响光滩土壤TS含量的关键影响因子, 此土层土壤含水量大于表层, 可溶性盐达最大值, 对土壤TS含量影响显著.互花米草入侵后10月土壤AS含量在0~10 cm深度最高(图 3), 原因可能与表面互花米草凋落物的分解有关, 有机质为影响互花米草潮滩土壤AS含量的关键因子(表 10), 该层有机质也处于垂直分布的最高值, 水热条件相对较好, 加速了有机质的分解.另外, 在12月, 互花米草湿地和光滩湿地0~10 cm土壤TS、AS含量均明显降低, 原因可能在于研究区12月夜间温度有时在-2 ℃以下, 可能出现退潮后土壤表层结冰现象, 冰融作用使表层土壤养分从表层向下迁移, 而此时2种湿地土壤表层可溶性盐含量亦明显降低.2种湿地类型均不同程度受到蟹类等动物的扰动, 由于蟹穴陷阱效应可能会使更多潮汐带来的SO42-富集, 使硫酸盐还原途径的优势升高(王琰等, 2017), 促使硫累积, 使之垂直波动均较小.不同时期, 互花米草潮滩各土层AS含量均显著大于光滩, 原因可能是互花米草入侵后, 根生物量大, 根际分泌物增多, 加之发达根系通气组织可将氧气带至深层土壤, 使得土壤矿化能力增强, 同时根际某些生物活性较强微生物的富集亦可促进硫矿化(李家兵等, 2016).
5 结论(Conclusions)1) 总体来看, 互花米草入侵后湿地土壤TS和AS平均含量(1.53、0.75 g·kg-1)明显高于入侵前的光滩湿地(1.13、0.53 g·kg-1), 其中, TS含量在10月增幅最大, 而AS含量在12月增幅最大.互花米草的入侵增加了土壤硫的时空变异性, 其入侵后不同月份土壤硫含量垂直变异性总体大于光滩湿地.
2) 互花米草各器官TS含量平均值表现为6月>10月>12月, 营养生长期(6月)含量最高(3.15 g·kg-1).植物生物量、硫累积量、AFs均与土壤AS有显著的相关性(p < 0.05), 互花米草对土壤硫的吸收与利用会影响土壤硫的分布.
3) 2种类型湿地土壤TS与AS含量之间均显著相关.互花米草湿地土壤AS含量的主要影响因子为土壤EC和有机质, 而土壤含水量是影响光滩湿地土壤TS、AS的关键因素.互花米草入侵后导致湿地pH、EC、有机质等理化性质的改变, 加之受互花米草特有的生长特性(硫累积特性)的影响, 使得湿地土壤硫循环更加复杂.
鲍士旦. 2000. 土壤农化分析[M]. 北京: 中国农业出版社: 159–161.
|
Cambardella C A, Moorman T B, Parkin T B, et al. 1994. Field-scale variability of soil properties in central iowa soils[J]. Soil Science Society of America Journal, 58(5): 1501–1511.
DOI:10.2136/sssaj1994.03615995005800050033x
|
Carlson P R J, Forrest J. 1982. Uptake of dissolved sulfide by Spartina alterniflora:Evidence from natural sulfur isotope abundance ratios[J]. Science, 216(4546): 633–635.
DOI:10.1126/science.216.4546.633
|
Chacón N, Herrera I, Flores S, et al. 2009. Chemical, physical, and biochemical soil properties and plant roots as affected by native and exotic plants in Neotropical arid zones[J]. Biology & Fertility of Soils, 45(3): 321–328.
|
邓自发, 安树青, 智颖飙, 等. 2006. 外来种互花米草入侵模式与爆发机制[J]. 生态学报, 2006, 26(8): 2678–2686.
DOI:10.3321/j.issn:1000-0933.2006.08.034 |
Ehrenfeld J G. 2003. Effects of exotic plant invasions on soil nutrient cycling processes[J]. Ecosystems, 6(6): 503–523.
DOI:10.1007/s10021-002-0151-3
|
Ferreira T O, Otero X L, Vidal-Torrado P, et al. 2007. Effects of bioturbation by root and crab activity on iron and sulfur biogeochemistry in mangrove substrate[J]. Geoderma, 142(1): 36–46.
|
郭雪莲, 吕宪国, 郗敏. 2007. 植物在湿地养分循环中的作用[J]. 生态学杂志, 2007, 26(10): 1628–1633.
|
郝庆菊, 王起超, 王跃思. 2003. 三江平原典型湿地的土壤植物总硫及水中硫酸盐的分布特征[J]. 生态环境学报, 2003, 12(4): 401–404.
DOI:10.3969/j.issn.1674-5906.2003.04.006 |
何涛, 孙志高, 李家兵, 等. 2016. 闽江河口不同淹水环境下典型湿地植物-土壤系统全硫含量空间分布特征[J]. 水土保持学报, 2016, 30(5): 246–254.
|
胡楚琦, 刘金珂, 王天弘, 等. 2015. 三种盐胁迫对互花米草和芦苇光合作用的影响[J]. 植物生态学报, 2015, 39(1): 92–103.
|
Jiang L F, Luo Y Q, Chen J K, et al. 2009. Ecophysiological characteristics of invasive Spartina alterniflora and native species in salt marshes of Yangtze River estuary, China[J]. Estuarine Coastal & Shelf Science, 81(1): 74–82.
|
Krairapanond N, Delaune R D, Patrick W H. 1991. Seasonal distribution of sulfur fractions in Louisiana salt marsh soils[J]. Estuaries, 14(1): 17–28.
DOI:10.2307/1351978
|
Lamers L P M, Govers L L, Janssen I C J M, et al. 2013. Sulfide as a soil phytotoxin-a review[J]. Front Plant Sci, 4(9): 268.
|
李家兵, 张秋婷, 张丽烟, 等. 2016. 闽江河口春季互花米草入侵过程对短叶茳芏沼泽土壤碳氮分布特征的影响[J]. 生态学报, 2016, 36(12): 3628–3638.
|
Lindsay W L. 1979. Chemical equilibria in soils[M]. Hoboken: Wiley.
|
林慧娜, 傅娇艳, 吴浩, 等. 2009. 中国主要红树林湿地沉积物中硫的分布特征及影响因素[J]. 海洋科学, 2009, 33(12): 79–82.
|
刘春悦, 张树清, 江红星, 等. 2009. 江苏盐城滨海湿地外来种互花米草的时空动态及景观格局[J]. 应用生态学报, 2009, 20(4): 901–908.
|
刘光崧. 1996. 土壤理化分析与剖面描述[M]. 北京: 中国标准出版社: 39–58.
|
李新华, 郭洪海, 杨丽萍, 等. 2014. 黄河三角洲翅碱篷湿地硫化氢和羰基硫排放动态研究[J]. 环境科学, 2014, 35(2): 786–791.
|
李新华, 刘景双, 孙志高, 等. 2007a. 三江平原小叶章湿地生态系统硫的生物地球化学循环[J]. 生态学报, 2007a, 27(6): 2199–2207.
|
李新华, 刘景双, 孙志高. 2007b. 三江平原小叶樟湿地植物-土壤系统中硫素分布及季节变化动态[J]. 土壤通报, 2007b, 38(1): 85–88.
|
李新华, 孙志高, 孙文广, 等. 2015. 黄河三角洲潮滩湿地系统二甲基硫排放通量的时空变化[J]. 环境科学学报, 2015, 35(12): 3947–3955.
|
李新华.2007.三江平原小叶章湿地土壤硫的形态分布和转化过程研究[D].长春: 中国科学院研究生院
|
Lu Q, Bai J, Gao Z, et al. 2016a. Effects of water level and salinity on total sulfur contents in salt marsh soils of the Yellow River Delta, China[J]. Wetlands, 36(1): 137–143.
|
Lu Q, Bai J, Fang H, et al. 2016b. Spatial and seasonal distributions of soil sulfur in two marsh wetlands with different flooding frequencies of the Yellow River Delta, China[J]. Ecological Engineering, 96(96): 63–71.
|
陆景陵. 2003. 植物营养学[M]. 北京: 中国农业大学出版社: 72–76.
|
Madureira M J, Vale C, Gonçalves M L S. 1997. Effect of plants on sulphur geochemistry in the Tagus salt-marshes sediments[J]. Marine Chemistry, 58(1): 27–37.
|
Mccary M A, Mores R, Farfan M A, et al. 2016. Invasive plants have different effects on trophic structure of green and brown food webs in terrestrial ecosystems:a meta-analysis[J]. Ecology Letters, 19(3): 328.
|
苗萍, 谢文霞, 于德爽, 等. 2017. 胶州湾互花米草湿地氮、磷元素的垂直分布及季节变化[J]. 应用生态学报, 2017, 28(5): 1533–1540.
|
牛书丽, 蒋高明, 李永庚. 2004. C3与C4植物的环境调控[J]. 生态学报, 2004, 24(2): 308–314.
|
佩奇A L, 米勒R H. 1991. 土壤分析法[M]. 北京: 中国农业科技出版社: 353.
|
Perkins E J, Streever W J, Davis E, et al. 2002. Development of amplified fragment length polymorphism markers for Spartina alterniflora[J]. Aquatic Botany, 74(1): 85–95.
DOI:10.1016/S0304-3770(02)00046-3
|
Richards C L, Pennings S C, Donovan L A. 2005. Habitat range and phenotypic variation in salt marsh plants[J]. Plant Ecology, 176(2): 263–273.
|
Seliskar D M, Gallagher J L, Burdick D M, et al. 2002. The regulation of ecosystem functions by ecotypic variation in the dominant plant:a Spartina alterniflora salt-marsh case study[J]. Journal of Ecology, 90(1): 1–11.
DOI:10.1046/j.0022-0477.2001.00632.x
|
Seliskar D M, Smart K E, Higashikubo B T, et al. 2004. Seedling sulfide sensitivity among plant species colonizing Phragmaites-infested wetlands[J]. Wetlands, 24(2): 426–433.
|
单孝全, 陈斌. 1991. 土壤和河流沉积物中硫的形态分析[J]. 环境科学学报, 1991, 11(2): 172–177.
DOI:10.3321/j.issn:0253-2468.1991.02.007 |
Simas T, Nunes J P, Ferreira J G. 2001. Effects of global climate change on coastal salt marshes[J]. Ecological Modelling, 139(1): 1–15.
|
Stribling J M, Cornwell J C. 2001. Nitrogen, phosphorus, and sulfur dynamics in a low salinity marsh system dominated by Spartina alterniflora[J]. Wetlands, 21(4): 629–638.
|
Sun Z G, Mou X J, Song H L, et al. 2013. Sulfur biological cycle of the different Suaeda salsa marshes in the intertidal zone of the Yellow River estuary, China[J]. Ecological Engineering, 53(2): 153–164.
|
王琰, 童春富. 2017. 长江口芦苇和互花米草盐沼湿地蟹类洞穴分布特征及主要影响因子[J]. 生态学报, 2017, 37(16): 5504–5513.
|
夏露.2015.入侵种互花米草(Spartina alterniflora L.)遗传生态及硫适应[D].南京: 南京大学
|
谢文霞, 朱鲲杰, 崔育倩, 等. 2014. 胶州湾河口湿地土壤有机碳及氮含量空间分布特征研究[J]. 草业学报, 2014, 23(6): 54–60.
|
许秀丽, 张奇, 李云良, 等. 2014. 鄱阳湖典型洲滩湿地土壤含水量和地下水位年内变化特征[J]. 湖泊科学, 2014, 26(2): 260–268.
|
Yu X Q, Yang J, Liu L M, et al. 2014. Effects of Spartina alterniflora invasion on biogenic elements in a subtropical coastal mangrove wetland[J]. Environmental Science & Pollution Research, 22(4): 3107–3115.
|
于君宝, 褚磊, 宁凯, 等. 2014. 黄河三角洲滨海湿地土壤硫含量分布特征[J]. 湿地科学, 2014, 12(5): 559–565.
|
曾从盛, 王维奇, 翟继红. 2010. 闽江河口不同淹水频率下湿地土壤全硫和有效硫分布特征[J]. 水土保持学报, 2010, 24(6): 246–250.
|
Zhou C F, An S Q, Deng Z, et al. 2009. Sulfur storage changed by exotic Spartina alterniflora in coastal saltmarshes of China[J]. Ecological Engineering, 35(4): 536–543.
DOI:10.1016/j.ecoleng.2008.01.004
|
Zhou J, Wu Y, Kang Q, et al. 2007. Spatial variations of carbon, nitrogen, phosphorous and sulfur in the salt marsh sediments of the Yangtze Estuary in China[J]. Estuarine Coastal and Shelf Science, 71(1): 47–59.
|
訾园园, 郗敏, 孔范龙, 等. 2016. 胶州湾滨海湿地土壤有机碳时空分布及储量[J]. 应用生态学报, 2016, 27(7): 2075–2083.
|