 2019, Vol. 39
2019, Vol. 39



2. 中国地质大学(北京)中国地质科学院, 北京 100083;
3. 合肥工业大学资源与环境工程学院, 合肥 230009;
4. 河北省地下水污染机理与修复重点实验室, 石家庄 050061;
5. 北京未名清源环保科技有限公司, 北京 100080
2. Chinese Academy of Geological Sciences, China University of Geosciences in Beijing, Beijing 100083;
3. School of Resources and Enviromental Engineering, HeFei University of Technology, Hefei 230009;
4. Key Laboratory of Groundwater Remediation of Hebei Province, Shijiazhuang 050061;
5. Beijing Weiming Qingyuan Environmental Protection Technology Co., Ltd., Beijing 100080
20世纪80年代前, 水文地球化学以天然岩石水化学作用研究为主, 随着地下水污染尤其是有机污染的增多, 污染有机碳微生物代谢与天然岩石水化学作用交互的微生物地球化学作用研究成为地下水污染机理研究的新方向, 其关键科学问题之一是如何识别无机碳(DIC)的碳循环过程(沈照理等, 2012).
普遍认为DIC变化是微生物代谢及碳酸盐岩水化学溶解/沉淀的结果.具体有以下4个作用:①异养微生物代谢:有机污染物为异养微生物提供碳源, 作为电子供体与O2、NO3-、Fe3+、Mn4+、SO42-等电子受体发生氧化还原反应, 有机污染物代谢为CO2, DIC升高(Bolliger et al., 1999); ②碳酸盐岩水化学溶解:代谢过程一般形成有机酸与CO2, 共同增加酸度侵蚀溶解碳酸盐岩, DIC升高(韩贵琳等, 2005); ③碳酸盐岩水化学沉淀:DIC增多, 或CO2分压减小, 或钙镁等阳离子增多, 使碳酸盐岩溶解过饱和, 形成DIC沉淀(潘根兴, 1999), 该过程DIC可升高可降低.④自养微生物代谢:自养微生物以DIC作为主要碳源(袁红朝等, 2011), 将DIC代谢为有机碳(OC), DIC降低(刘彦等, 2010), 在此过程中, 微生物创造会形成碱性微环境, 在菌体表面成核等作用下, 使DIC与Ca2+、Mg2+等阳离子形成沉淀, 导致DIC降低(周小萍等, 2013; 王红梅等, 2013; Xu et al., 2017).
目前, 大多数有机污染地下水系统的DIC呈高异常特征.Suarez等(2002)在美国、Marić等(2018)在塞尔维亚、苏小四等(2011)、焦珣等(2012)在我国东北等地的研究均显示有机污染地下水中DIC高异常.然而, 华北平原某石油污染场地监测发现, 地下水污染区DIC在监测期间(2年共5次监测)均呈现明显低异常特征.海洋、地表水、土壤研究发现, 存在自养微生物代谢及诱导产生碳酸盐岩沉淀现象(Dittrich et al., 2004; 袁红朝等, 2011).地下水中也发现存在有大量的化能自养微生物(Alfreider et al., 2012).但自养微生物能否导致石油污染地下水DIC低异常现象仍没有明确答案.
为此, 在水文地球化学分析和16S rRNA基因高通量测序基础上, 结合含水层结构及流场特征, 剖析了地下水的水化学和微生物两方面作用, 辨识了地下水中DIC低异常的主导因素, 揭示了生物地球化学作用的机理, 探析该石油污染地下水DIC低异常成因, 在此基础上探讨了DIC的空间变异规律.
2 材料与方法(Materials and methods) 2.1 场地概况研究区位于华北平原某待搬迁石油污染场地, 其油罐已有40余年, 泄漏所导致地下水受到石油类污染严重.在油罐泄漏区及其周边布设了11口监测取样井(图 1).
 |
| 图 1 污染泄露区域、监测井位置及流场示意图 Fig. 1 Map of leakage area, monitoring wells and groundwater flow |
该地区地下水含水层结构如图 2所示, 以多层砂砾石和少数砂层为主, 下部存在粉质粘土层, 可视为下部的隔水层.地下水位埋深21.7~24.0 m, 存在0.4~2.0 m厚的粉质粘土层, 可视为上部的隔水层, 稳定水头高于含水层顶板, 可视以上分区含水层为承压含水层.而下游的MW17和MW6钻孔资料揭示, 在地下水水位附近, 未发现可作为上部隔水层的土层, 视为潜水.
 |
| 图 2 污染场地含水层剖面示意图 Fig. 2 Schematic diagram of aquifer profile in the contaminated site |
从地下水流场(图 1)可知, 地下水在流经污染源后, 流向由“北西-南东”变为“西南-东北”, 偏转约90°, 结合含水层特征, 下游正东方向(MW6)和东北方向(MW17)无粉质粘土隔水层, 而污染源周边及下游东南方向(MW10)有较厚粉质粘土的上部隔水层, 地下水相对滞流.
2.2 样品的采集与测试根据前期布井方案(张敏等, 2017), 于2016年8月用贝勒管采集样品11个.为了保证所采样品的代表性, 采样前使用便携式水质仪测定井水水质参数, 当3次测定参数值变化范围达到:水温(T)±1 ℃、pH ±0.2、氧化还原电位(ORP)±20 mV、电导率(EC)±3%及溶解氧(DO)±10%或±0.2时, 采集水样, 置于4 ℃便携式冰箱中保存并送回实验室进行测试.根据中国生活饮用水卫生标准检验方法(GBT 5750.6.2.1 —2006), 测试化学需氧量(COD)、硝酸盐(NO3-)、硫酸盐(SO42-)、重碳酸盐(HCO3-)和钙(Ca2+)、镁(Mg2+)等离子浓度.利用气相色谱-质谱法测定了苯系物(BTEX)浓度.测试结果如表 1所示.
| 表 1 样品测试结果 Table 1 Determination of physicochemical parameters of different samples |
为了解地下水化学成分的变化原因, 对研究区的水样微生物种群结构进行了基于16S rRNA基因的测序分析.具体步骤为:1 L水样经孔径为0.22 μm的PTFE滤膜过滤采集DNA, 用EZNATM DNA提取试剂盒(OMEGA bio-tek, Norcross, GA, USA)提取DNA, 扩增其16S rRNA基因V4~V5区(Ning et al., 2018), 并在派森诺公司HiSeq 2500平台测序.测序数据进行质控过滤与拼接, 对优化序列的非重复序列(不含单序列), 按照97%相似性聚类进行可操作单元(Operational taxonomic unit, OTU)聚类, 在聚类过程中去除嵌合体, 得到OTU的代表序列(蔡萍萍等, 2018).利用相关分析法, 在美吉生物信息云平台(http://www.i-sanger.com/), 进行物种OTU与DIC、Ca2+、Mg2+、硬度的Spearman相关性分析, 推测形成该含水层DIC低异常的微生物作用.
3 结果与讨论(Results and discussion) 3.1 地下水污染分布及水化学类型地下水污染分布如图 3所示.苯系物浓度在污染源处浓度最大, 沿着地下水流动方向依次减小; COD最大值出现在MW17井, 浓度次高点位于PM4, 其它位置沿着地下水流动方向依次减小.苯系物为石油污染物很小一部分, 但其毒性大, 易挥发, 为重点关注污染物(张玉欣等, 2017).它具有易吸附、难迁移的特点(Basu et al., 2017), 因此在污染源处集中分布; COD可指示地下水石油类有机污染物程度(Wan et al., 2016), 它指示了所有污染物, 包括难迁移和易迁移物质.污染源和近下游是COD高值区, 污染延伸主导方向为东-东北方向.由此可知, 污染物的分布由水力作用主导, 同时与污染物的吸附降解等性质有关.
 |
| 图 3 地下水污染数值(COD和苯系物)分布图 Fig. 3 Contamination indexes distribution in groundwater |
Piper三线图(图 4)可看出, 从背景区到污染区的地下水类型由HCO3·Cl·SO4-Ca·Mg演变为HCO3·Cl-Ca·Mg.由于硫酸盐还原作用的存在, SO42-流经污染区后含量减小, 而HCO3-相对含量增大.除上游更远的(MW4和MW5)外, 其余样品的HCO3-含量均超过50%.该场地地下水阳离子中以Ca2+、Mg2+为主, 含量占比均超50%, 但相对于背景区, Ca2+、Mg2+比值有所减小而Na+、K+比值有所增加.
 |
| 图 4 地下水Piper三线图 Fig. 4 Piper Diagram of groundwater samples |
上游背景井MW4具有较高的DIC含量, 为170 mg·L-1; 紧邻污染源的上游PM7和OTAW4井DIC含量最低, 分别为65和101 mg·L-1; 沿着水流方向, 从PM7到污染源下游, DIC含量逐渐增加, MW17和MW6超过180 mg·L-1.
根据碳酸盐沉淀-溶解平衡(反应(1)), Ca2+、Mg2+含量的增加会导致DIC的减少.该场地的DIC与硬度分布一致(图 5b).因此, 该场地DIC的减少不是Ca2+、Mg2+增加引起的.
 |
| 图 5 地下水DIC、硬度、pH分布图 Fig. 5 Distribution of groundwater DIC, hardness and pH |
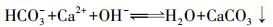
|
(1) |
Ca2+、Mg2+等阳离子被地下水微生物利用的能力有限, 且不会形成气体, 只能以矿化而沉淀的形式离开地下水体系, 且Ca2+、Mg2+较易形成碳酸盐沉淀.一般认为Ca2+、Mg2+的减少指示碳酸盐的沉淀.从该含水层的DIC分布图(图 5a)可以看出, 背景区及污染下游区具有较高含量的DIC, 污染源及周边区域DIC含量较低.这恰好与硬度具有相似地分布.
减小CO2分压可使DIC形成沉淀(反应(2)).据各井pH和α(CO2)(CO2与DIC物质的量之比)关系, 拟合封闭体系碳酸平衡理论模型方程, R2=0.9977, 指示该含水层可视为封闭体系.封闭体系中, 没有CO2的散失, 不造成纯水化学导致的碳酸盐沉淀.
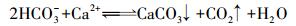
|
(2) |
pH值(图 5c)与DIC和硬度具有相反的分布.碱性环境容易导致碳酸盐沉淀的形成(于炳松等, 2006).由此推测, 该场地地下水中的DIC和Ca2+、Mg2+等离子会形成碳酸盐沉淀而使DIC和Ca2+、Mg2+减小.
3.3 微生物代谢对DIC空间分布的影响① 异养微生物代谢产DIC污染区域含水层有较多的石油有机质, 污染的长期存在会聚集以有机物为电子供体而消耗硫酸盐和硝酸盐等电子受体的微生物, 造成SO42-和NO3-浓度的减小(刘新华等, 1997; 张敏等, 2017).在此过程中, 污染物的降解会释放CO2, 溶于水形成HCO3-, 造成HCO3-浓度在一定程度上的升高(Marić et al., 2018).相比于背景区, 该场地污染区域所有电子受体的浓度在污染部位明显减少(张敏等, 2017), Spearman热图上多种微生物(用OTU代表)与DIC显著正相关, 指示有机碳转化为DIC.
一般认为, 有机物的生物降解会导致地下水中的DIC升高.根据勒沙特列原理, DIC浓度的增加, 导致Ca2+、Mg2+的沉淀, 但并不会使水中DIC浓度降低.
② 自养微生物代谢消耗DIC从Spearman关联热图(图 6)上可以看出, DIC与OTU16、OTU53、OTU482和OTU541显著负相关(p < 0.05).OTU16对应Pseudomonas菌属, 为兼性营养菌(Blackmore et al., 1968), 某些菌种具有自养反硝化功能(黄俊等, 2015), 某些菌种能促进碳酸盐沉淀(Hammes et al., 2002, Sarkisova et al., 2005), 还有些菌可利用氢气作为能源, 通过碳基歧化酶、磷酸烯醇丙酮酸羧酶和苹果酸酶, 把CO2转化为多种氨基酸、糖类和酯类(Donawa et al., 1973); OTU53对应Pseudoxanthomonas菌属, 可以兼性营养反硝化(Xu et al., 2015), 利用氢气作为能源, 把DIC转化为有机碳; OTU482在NCBI数据库中还未有相匹配的菌, 与其最大相似率(91%)的未培养微生物(GenBank: KJ149220.1)与碳酸盐有关(Postec et al., 2015).OTU541对应未培养的Sedimentibacter菌属, 该菌能够利用有机物产生氢气(贾璇等, 2016), 为氢营养型微生物生长提供营养.
 |
| 图 6 OTU与DIC、Ca2+、Mg2+、硬度的Spearman相关性分析 Fig. 6 Spearman correlation analysis between OTU and DIC, Ca2+, Mg2+, hardness |
Ca2+和Mg2+与OTU558显著负相关(p < 0.05), Mg2+与OTU541显著负相关(p < 0.05).OTU558对应Polynucleobacter属, 可能与CO2的非光合固定有关(Casamayor et al., 2012), 可吸收CO2 (Negandhi et al., 2014).OTU541同时与DIC显著负相关.
从图中可以看出, 大部分与DIC负相关微生物同时与Ca2+、Mg2+和硬度负相关.但在该场地有种特殊的微生物OTU333, 其与Ca2+、Mg2+负相关而与DIC弱正相关, 其大约占整个场地的OTU的14%.在污染源部位的OTAW4、MW3和PM7中占比分别为32%、36%和20%, 这些井恰好是DIC和Ca2+、Mg2+含量的最低点.污染最严重MW17井的OTU333丰度为38%, 位于污染侧翼MW13井的OTU333丰度为11%, 这两井有较高的DIC和较低的Ca2+、Mg2+含量.其余井的OTU333的丰度小于10%, 这些井DIC和Ca2+、Mg2+含量均较高.OTU333对应的微生物菌种为包括Hydrogenophaga defluvii strain在内的多种Hydrogenophaga菌属, 碱基均为100%匹配.Hydrogenophaga菌能够利用H2和DIC自养(Kämpfer et al., 2005), 是一种很重要的固碳微生物(Liu et al., 2015).这就解释了除MW17外, OTU333丰度高的样品具有较小的DIC.MW17污染物浓度较高, 其有机物降解产生DIC的速率大于Hydrogenophaga对DIC消耗的速率, 造成DIC积累的现象.根据反应方程式(2)和(6), 在自养过程中, 伴随着DIC的消耗, 会形成碳酸钙镁沉淀, 造成地下水中钙镁离子的减小.此外, Hydrogenophaga在氢化酶(hydrogenase)作用下产生氢气(Yoon et al., 2009), 供给自养微生物生长.
本研究还未对古菌进行分析, 不能直接判断该场地地下水是否有产甲烷作用.但通过前期研究表明, 该场地极有可能发生着产甲烷作用(张敏等, 2017).产甲烷作用也是消耗DIC的过程.有些微生物OTU虽然与DIC没有显著相关, 但也可能与DIC循环有关, 在此不再做深入讨论.
3.4 低异常成因探析根据上述分析推测, 该场地地下水DIC减少主要有两种作用:一种是微生物把DIC转化为有机碳(刘彦等, 2010); 另一种是地下水的碱性环境促使碳酸盐的沉淀.
第一种作用的微生物以化能自养微生物为主, 它们利用DIC合成细胞有机质.以噬氢自养类微生物为例, 发生如下反应.
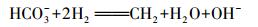
|
(3) |
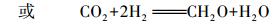
|
(4) |
该反应中, 每消耗1 mol DIC, 形成1 mol有机碳.
研究表明, 石油等有机污染场地的污染源部位会发生产甲烷作用(Marić et al., 2018), 其中, 噬氢产甲烷作用可表示为:
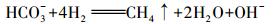
|
(5) |
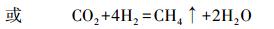
|
(6) |
该反应中, 同样每消耗1 mol DIC, 形成1 mol有机碳.
根据反应(1)、反应(3)和(5)生成的OH可促使碳酸盐沉淀的形成.反应(4)和(6)消耗CO2, 促进反应(2)向沉淀方向进行.对于某些微生物, 其代谢可产生的OH-, 在细胞周围形成碳酸盐沉淀微环境, 在几乎不改变整个水化学成分的条件下完成碳酸盐沉淀(王红梅等, 2013).若OH生成速率大于沉淀速率, 则会造成地下水OH-的积累, 导致地下水pH升高, 诱导第二种作用的发生.
结合场地微生物分析, 推测该场地DIC低异常主要原因为氢营养型化能自养微生物代谢的结果.该过程分为3步:第一步为石油类有机物在氢化酶的作用下产氢气过程, 主要可能参与的微生物为Hydrogenophaga和Sedimentibacter菌属; 第二步为DIC在固碳微生物和产甲烷微生物作用下, 利用第一步产生的H2, 发生反应(3)~(6), 合成有机碳(包括甲烷)并产生OH-的过程, 主要可能参与的微生物为Hydrogenophaga、Pseudomonas、Pseudoxanthomonas、Polynucleobacter及产甲烷菌; 第三步为DIC的沉淀作用, 利用第二步产生的OH-, 以反应(1)的方式, 促使碳酸盐的形成.
4 结论(Conclusions)1) 该石油污染含水层中的天然地下水化学类型以HCO3·Cl·SO4-Ca·Mg为主, 流经污染区后演变为HCO3·Cl-Ca·Mg型.
2) 该含水层碳酸平衡符合封闭体系模型, 无CO2逸散导致的DIC沉淀.
3) 该含水层不仅存在着异养微生物, 还可能存在大量自养微生物, 自养微生物代谢导致了DIC的减少并形成钙镁的碳酸盐沉淀.因此, 该场地地下水DIC低异常可能与该区域地下水具有较高浓度的钙镁离子和较高活性的自养微生物代谢有关.
Alfreider A, Schirmer M, Vogt C. 2012. Diversity and expression of different forms of RubisCO genes in polluted groundwater under different redox conditions[J]. FEMS Microbiology Ecology, 79(3): 649–660.
DOI:10.1111/fem.2012.79.issue-3
|
Basu S, Yadav B K, Mathur S. 2017. Modeling Systems and Processes in Wetlands: A Case Study of Engineered Bioremediation of BTEX-Contaminated Water in Treatment Wetlands[M]//Prusty B A K, Chandra R, Azeez P A, Wetland Science: Perspectives From South Asia. Springer India; New Delhi: 463-488
|
Blackmore M A, Quayle J R. 1968. Choice between autotrophy and heterotrophy in Pseudomonas oxalaticus. Growth in mixed substrates[J]. Biochemical Journal, 107(5): 705.
DOI:10.1042/bj1070705
|
Bolliger C, HöHener P, Hunkeler D, et al. 1999. Intrinsic bioremediation of a petroleum hydrocarbon-contaminated aquifer and assessment of mineralization based on stable carbon isotopes[J]. Biodegradation, 10(3): 201–217.
DOI:10.1023/A:1008375213687
|
Casamayor E O, Llir S M, Picazo A, et al. 2012. Contribution of deep dark fixation processes to overall CO2 incorporation and large vertical changes of microbial populations in stratified karstic lakes[J]. Aquatic Sciences, 74(1): 61–75.
DOI:10.1007/s00027-011-0196-5
|
蔡萍萍, 宁卓, 何泽, 等. 2018. 采油井场土壤微生物群落结构分布[J]. 环境科学, 2018, 39(7): 3329–3338.
|
Dittrich M, Obst M. 2004. Are picoplankton responsible for calcite precipitation in lakes?[J]. Ambio, 33(8): 559–564.
DOI:10.1579/0044-7447-33.8.559
|
Donawa A L, Ishaque M, Aleem M I. 1973. CO2 fixation and metabolic control in Pseudomonas saccharophila[J]. Canadian Journal of Microbiology, 19: 1243–1250.
DOI:10.1139/m73-201
|
Hammes F, Verstraete W. 2002. Key roles of pH and calcium metabolism in microbial carbonate precipitation[J]. Reviews in Environmental Science and Biotechnology, 1(1): 3–7.
|
韩贵琳, 刘丛强. 2005. 贵州喀斯特地区河流的研究——碳酸盐岩溶解控制的水文地球化学特征[J]. 地球科学进展, 2005, 20(4): 394–406.
DOI:10.3321/j.issn:1001-8166.2005.04.004 |
黄俊, 张诗颖, 王翻翻, 等. 2015. 同步脱氮除硫菌的筛选鉴定及其生长特性研究[J]. 生物技术通报, 2015, 31(10): 184–190.
|
贾璇, 任连海, 李鸣晓, 等. 2016. 芦苇秸秆厌氧联产氢气甲烷过程中细菌群落演替规律[J]. 农业工程学报, 2016, 32(4): 199–204.
|
焦珣, 苏小四, 吕航. 2012. 某石油类污染场地地下水石油烃生物降解的地球化学证据[J]. 地质科学, 2012, 47(2): 499–506.
DOI:10.3969/j.issn.0563-5020.2012.02.017 |
Kämpfer P, Schulze R, Jäckel U, et al. 2005. Hydrogenophaga defluvii sp. nov. and Hydrogenophaga atypica sp. nov., isolated from activated sludge[J]. International Journal of Systematic and Evolutionary Microbiology, 55(1): 341–344.
DOI:10.1099/ijs.0.03041-0
|
Liu J F, Sun X B, Yang G C, et al. 2015. Analysis of microbial communities in the oil reservoir subjected to CO2-flooding by using functional genes as molecular biomarkers for microbial CO2 sequestration[J]. Frontiers in Microbiology, 6(236): 236.
|
刘新华, 沈照理, 傅家谟. 1997. 地下水油类污染的水文地球化学指标——以山东省淄博市某地下水水源地为例[J]. 沉积学报, 1997, 15(2): 236–240.
|
刘彦, 张金流, 何媛媛, 等. 2010. 单生卵囊藻对DIC的利用及其对CaCO3沉积影响的研究[J]. 地球化学, 2010, 39(2): 191–196.
|
Marić N, Matić I, Papić P, et al. 2018. Natural attenuation of petroleum hydrocarbons-a study of biodegradation effects in groundwater (Vitanovac, Serbia)[J]. Environmental Monitoring and Assessment, 190(2): 89.
DOI:10.1007/s10661-018-6462-4
|
Negandhi K, Laurion I, Lovejoy C. 2014. Bacterial communities and greenhouse gas emissions of shallow ponds in the High Arctic[J]. Polar Biology, 37(11): 1669–1683.
DOI:10.1007/s00300-014-1555-1
|
Ning Z, Zhang M, He Z, et al. 2018. Spatial pattern of bacterial community diversity formed in different groundwater field corresponding to electron donors and acceptors distributions at a petroleum-contaminated site[J]. Water, 10: 842.
DOI:10.3390/w10070842
|
Postec A, Qu M, Neur M, et al. 2015. Microbial diversity in a submarine carbonate edifice from the serpentinizing hydrothermal system of the Prony Bay (New Caledonia) over a 6-year period[J]. Frontiers in Microbiology, 6(6): 857.
|
潘根兴. 1999. 中国干旱性地区土壤发生性碳酸盐及其在陆地系统碳转移上的意义[J]. 南京农业大学学报, 1999, 22(1): 51–57.
|
Sarkisova S, Patrauchan M A, Berglund D, et al. 2005. Calcium-induced virulence factors associated with the extracellular matrix of mucoid pseudomonas aeruginosa biofilms[J]. Journal of Bacteriology, 187(13): 4327–4337.
DOI:10.1128/JB.187.13.4327-4337.2005
|
沈照理, 王焰新, 郭华明. 2012. 水-岩相互作用研究的机遇与挑战[J]. 地球科学-中国地质大学学报, 2012, 37(2): 207–219.
|
苏小四, 吕航, 张文静, 等. 2011. 某石油污染场地地下水石油烃生物降解的(13)C、(34)S同位素证据[J]. 吉林大学学报(地), 2011, 41(3): 847–854.
|
Suarez M P, Rifai H S. 2002. Evaluation of BTEX remediation by natural attenuation at a coastal facility[J]. Groundwater Monitoring & Remediation, 22(1): 62–77.
|
Wan J, Gu J, Zhao Q, et al. 2016. COD capture:a feasible option towards energy self-sufficient domestic wastewater treatment[J]. Scientific Reports, 6: 25054.
DOI:10.1038/srep25054
|
王红梅, 吴晓萍, 邱轩, 等. 2013. 微生物成因的碳酸盐矿物研究进展[J]. 微生物学通报, 2013, 40(1): 180–189.
|
Xu G, Peng J, Feng C, et al. 2015. Evaluation of simultaneous autotrophic and heterotrophic denitrification processes and bacterial community structure analysis[J]. Applied Microbiology and Biotechnology, 99(15): 6527–6536.
DOI:10.1007/s00253-015-6532-2
|
Xu Q, Zhang C, Li F, et al. 2017. Arthrobacter Sp. Strain MF-2 induces high-mg calcite formation:Mechanism and implications for carbon fixation[J]. Geomicrobiology Journal, 34(2): 157–165.
DOI:10.1080/01490451.2016.1155002
|
Yoon K S, Sakai Y, Tsukada N, et al. 2009. Purification and biochemical characterization of a membrane-bound[NiFe]-hydrogenase from a hydrogen-oxidizing, lithotrophic bacterium, Hydrogenophaga sp. AH-24[J]. FEMS Microbiology Letters, 290(1): 114–120.
DOI:10.1111/fml.2008.290.issue-1
|
于炳松, 赖兴运. 2006. 成岩作用中的地下水碳酸体系与方解石溶解度[J]. 沉积学报, 2006, 24(5): 627–635.
DOI:10.3969/j.issn.1000-0550.2006.05.002 |
袁红朝, 秦红灵, 刘守龙, 等. 2011. 固碳微生物分子生态学研究[J]. 中国农业科学, 2011, 44(14): 2951–2958.
DOI:10.3864/j.issn.0578-1752.2011.14.012 |
张敏, 王森杰, 陈素云, 等. 2017. 地下水苯系物微生物降解及其碳同位素标记[J]. 水文地质工程地质, 2017, 44(2): 129–136.
|
张玉欣, 安俊琳, 王健宇, 等. 2017. 南京北郊大气BTEX变化特征和健康风险评估[J]. 环境科学, 2017, 38(2): 453–460.
|
周小萍, 蓝家程, 张笑微, 等. 2013. 岩溶溪流的脱气作用及碳酸钙沉积——以重庆市南川区柏树湾泉溪流为例[J]. 沉积学报, 2013, 31(6): 1014–1021.
|