 2019, Vol. 39
2019, Vol. 39



2. 南京农业大学资源与环境科学学院, 南京 210095
2. College of Resources and Environmental Sciences, Nanjing Agricultural University, Nanjing 210095
2016年, 国务院印发了《土壤污染防治行动计划》, 旨在加强我国土壤污染防治, 逐步改善土壤环境质量, 确保农产品安全和人群健康.多环芳烃(PAHs)是土壤环境中普遍存在的一类持久性有机污染物(POPs), 具有高生物富集、致毒、致癌和难降解等特点(Menzie et al., 1992; Wang et al., 2012).人类生产活动中产生的PAHs可以通过污水灌溉、大气沉降、石油泄漏、农业废物燃烧和化肥施用等多种途径进入农田生态系统(Samsøe-Petersen et al., 2002; Cai et al., 2017).PAHs进入农田土壤环境后, 可以被农作物根部吸收、积累并转运至茎叶, 进而沿食物网传递危害野生物种和人群健康(Kim et al., 2013;陈保冬等, 2015; Chen et al., 2015).如何降低土壤和农作物PAHs污染、保护农业耕地、生产出绿色安全的农产品已经成为国内外关注的热点问题之一.
研究指出, 接种功能细菌不仅可以促进植物生长(Çakmakçi et al., 2006; Glick, 2014), 也能够降低植物PAHs污染(Arslan et al., 2017; Waigi et al., 2017).例如, Liu等(2014; 2017)从看麦娘(Alopecurus aequalis Sobol)体内获得一株具有菲降解特性的菌株Massilia sp. Pn2, 该菌能够重新定殖在黑麦草(Lolium multiflorum Lam)和小麦(Triticum aestivum L. cv. Yangmai-16)体内, 促进植物生长并降低植物体内菲含量.然而, 关于接种混合功能菌降低植物PAHs污染的作用机理仍不清楚(刘爽等, 2013;孙凯等, 2014; Liu et al., 2017).已有资料显示, 植物吸收PAHs后, 其体内过氧化物酶(POD)可以清除植物体内超氧自由基, 控制膜脂的过氧化作用并保护细胞膜的正常代谢(Kvesitadze et al., 2009;夏红霞等, 2013);而植物体内多酚氧化酶(PPO)能够催化氧化PAHs开环, 生成易降解的中间产物(Iwabuchi et al., 1998).由此可见, 植物体内POD和PPO活性大小可能影响其对PAHs的应答反应和代谢水平.能否通过接种功能菌改变植物体内POD和PPO活性, 调节植物体内PAHs的降解代谢?该想法值得深入探究.
菲和芘作为3环和4环PAHs的典型代表, 常被用于研究PAHs的微生物降解和植物代谢过程(Barnier et al., 2014; Zielińska et al., 2015).本研究选择实验室前期获得的两株分别具有高效菲和芘降解特性的Massilia sp. Pn2和Mycobacterium flavescens 033作为供试菌株, 通过批量降解试验方法, 探讨菌株Pn2和033降解菲和芘的基本动力学过程和规律;重点采用温室盆栽试验方法, 研究接种混合菌株Pn2和033对黑麦草体内PAHs含量及PPO和POD活性的影响.以期从植物体内PAHs含量及其PPO和POD活性方面阐明接种混合菌降低植物PAHs污染的作用机制, 为利用植物-功能菌的微生态调控技术规避作物PAHs污染风险和保障人群健康提供理论依据.
2 材料与方法(Materials and methods) 2.1 化学试剂和培养基菲和芘购自德国Fluka公司, 纯度>98%, 其理化性质如图 1所示.甲醇为色谱醇, 其它试剂为分析纯.利用丙酮制备浓度分别为4 mg·mL-1和2 mg·mL-1的菲和芘母液, 置于4 ℃冰箱保存备用.
 |
| 图 1 菲和芘理化性质 Fig. 1 Selected physicochemical properties of phenanthrene and pyrene in this study |
LB培养基:蛋白胨10.0 g·L-1, 酵母粉5.0 g·L-1, NaCl 10.0 g·L-1, pH=7.0.
无机盐培养基(MSM):(NH4)2SO4 1.50 g·L-1, K2HPO4·3H2O 1.91 g·L-1, KH2PO4 0.50 g·L-1, MgSO4·7H2O 0.20 g·L-1, 2 mL微量元素溶液(CoCl2·6H2O 0.1 mg·L-1, MnCl2·4H2O 0.425 mg·L-1, ZnCl2 0.05 mg·L-1, NiCl2·6H2O 0.01 mg·L-1, CuSO4·5H2O 0.015 mg·L-1, Na2MoO4·2H2O 0.01 mg·L-1, Na2SeO4·2H2O 0.01 mg·L-1), pH=7.0.
固体培养基:添加1.8%琼脂, 制备固体LB培养基和固体MSM.
霍格兰营养液:Ca(NO3)2·4H2O 945 mg·L-1, KNO3 506 mg·L-1, NH4NO3 80 mg·L-1, KH2PO4 136 mg·L-1, MgSO4 493 mg·L-1, 2.5 mL铁盐溶液(FeSO4·7H2O 2.78 g, EDTA·Na2 3.73 g), 500 mL去离子水(18.2 MΩ·cm), 5 mL微量元素溶液(KI 0.83 mg·L-1, H3BO3 6.2 mg·L-1, MnSO4 22.3 mg·L-1, ZnSO4 8.6 mg·L-1, Na2MoO4 0.25 mg·L-1, CuSO4 0.025 mg·L-1, CoCl2 0.025 mg·L-1, pH 6.0), pH=5.5.
2.2 供试植物和菌株选取生长旺盛、根系发达且脂肪含量高的黑麦草(Lolium multiflorum Lam)作为供试植物.本实验室前期从南京某PAHs污染区的健康植物看麦娘(Alopecurus aequalis)体内获得一株具有高效菲降解特性的革兰氏阴性菌株Massilia sp. Pn2, 该菌在含有50 mg·L-1芘的固体MSM上生长良好, 且能够定殖在黑麦草体内(Liu et al., 2014).芘降解菌株Mycobacterium flavescens 033购自美国菌种保藏中心, 属于革兰氏阳性菌,该菌对100 mg·L-1菲具有较强的耐受性, 并能够在植物体内定殖(Dean-Ross et al., 1996; Wang et al., 2017).分别采用含有高浓度菲和芘的MSM扩大培养菌株Pn2和033, 当菌株生长至对数期时, 将培养液置于高速离心机, 8000 r·min-1下离心5 min, 反复用无菌水清洗菌体表面吸附的PAHs后, 用灭菌的MSM制备菌悬液.
2.3 批量降解试验采用批量降解试验方法, 分别测定功能菌株Pn2和033对菲和芘的降解效率和基本动力学参数(Zhu et al., 2016).将0.5 mL菲和芘的丙酮母液加入到50 mL灭菌三角瓶中, 置于无菌操作台, 待丙酮挥发后, 向三角瓶中添加20 mL灭菌MSM, 分别获得以100 mg·L-1菲和50 mg·L-1芘作为碳源和能源的PAHs降解培养基.按照5%接种量分别在含有菲和芘的MSM中加入OD600=1.0、处于对数生长期的菌株Pn2(菌密度为1.26×107 CFU·mL-1)和033(菌密度为1.17×107 CFU·mL-1)菌悬液(菌悬液为灭菌的MSM), 置于30 ℃、150 r·min-1避光摇床培养, 定时取样, 测定培养液中细菌数量及PAHs残留浓度.设置接种灭活菌株处理组作为空白对照, 每组实验重复3次.
采用整瓶提取方法测定培养基中PAHs残留浓度.向培养基中加入等体积色谱甲醇, 超声萃取30 min, 8000 r·min-1高速离心后, 过0.22 μm滤膜, 高效液相色谱(HPLC, Shimadzu LC-20AT, Japan)测定溶液中PAHs残留浓度.HPLC设定参数:Inertsil ODS-SP-C18反相色谱柱(150 mm×4.6 mm, 5 μm), UV/Vis检测器(SPD-20A), 流动相为甲醇/水=90:10(V/V), 流速为1.0 mL·min-1, 柱温为40 ℃, 检测波长为245 nm, 进样量为20 μL.
2.4 温室盆栽试验采用温室盆栽试验方法, 研究接种混合菌株对植物体内PAHs含量及其PPO和POD活性的影响.以南京江宁区距离土壤表面0~20 cm农田黄棕壤作为供试土壤,土壤理化性质为:pH=6.02, 有机碳含量14.3 g·kg-1, 黏土含量24.7%, 沙含量67.9%.土壤样品风干后, 过20目筛, 用含有菲和芘的丙酮母液污染.将污染土壤置于黑暗室温老化30 d后, 测得土壤中菲和芘的含量分别为84.65和43.58 mg·kg-1.试验设置6种不同处理, 具体如表 1所示.黑麦草种子经75%乙醇表面消毒5 min后, 用去离子水清洗干净, 置于30 ℃恒温培养箱催芽48 h.每盆(350 g土壤)种植定量经过催芽的黑麦草种子, 待黑麦草生长至幼苗期进行间苗, 每盆留取10株黑麦草幼苗继续生长.人工气候箱温度设置为25/20 ℃(白昼), 湿度50%.培养过程中适量补充半强度的霍格兰营养液, 以保持土壤中养分含量.待黑麦草植株生长至10 cm高时, 采用灌根法, 在其根部土壤接种10 mL OD600=2.0的混合菌悬液(菌悬液为灭菌的半强度霍格兰营养液, 菌株Pn2菌密度为5.63×108 CFU·mL-1, 菌株033菌密度为4.29×108 CFU·mL-1), 培养30 d后, 收集黑麦草根和茎叶样品, 测定其鲜重和干重并分析PAHs含量.每组实验重复3次.
| 表 1 温室盆栽试验中6种不同处理的黄棕壤 Table 1 Yellow-brown soil of six different treatments in the greenhouse pot experiments |
土壤和黑麦草样品中菲和芘的提取和测定方法参照文献(Gao et al., 2004).具体操作方法如下:①黑麦草样品冷冻干燥后, 用研钵研磨粉碎, 称取一定量的黑麦草干重样品置于30 mL玻璃离心管中, 用30 mL的二氯甲烷和正己烷(1:1, V/V)溶液分3次, 每次10 mL超声萃取30 min(土壤样品每次超声萃取60 min);②将萃取液收集后过无水硫酸钠-硅胶柱净化, 再次用5 mL的二氯甲烷和正己烷(1:1, V/V)洗脱净化柱中残留PAHs;③洗脱液收集至旋转蒸发瓶, 40 ℃恒温浓缩至干, 甲醇定容到1 mL, 过0.22 μm孔径滤膜后, HPLC定量分析土壤/黑麦草样品中菲和芘残留浓度.
2.6 黑麦草体内PPO和POD活性测定采用Shah等(2001)研究方法, 测定黑麦草体内PPO和POD活性, 并略做改进.分别称取0.1 g黑麦草根和茎叶样品, 置于冰浴研钵中, 加入0.05 g聚乙烯吡咯烷酮(PVP)和3 mL pH=7.8的磷酸盐缓冲溶液(PBS), 充分研磨均匀.采用高速离心机在4 ℃、10000 r·min-1下离心10 min, 上清液为粗酶提取液.利用邻苯二酚显色法测定PPO活性, 向玻璃试管中快速添加2 mL 0.01 mol·L-1 PBS(pH=6.0)、1.0 mL 11 g·L-1邻苯二酚和40 μL粗酶液, 充分摇匀后, 置于30 ℃水浴锅中水浴10 min, 于420 nm波长下测定其吸光度值.利用愈创木酚显色法测定POD活性, 向玻璃试管中迅速加入2 mL 0.1 mol·L-1醋酸缓冲液(pH=5.0)、1.0 mL 2.5 g·L-1愈创木酚、0.1 mL 3% H2O2和20 μL粗酶液.将混合液摇匀后, 置于37 ℃水浴锅水浴15 min, 在470 nm波长下测定其吸光度值.每组实验设置5个重复.酶活性(R)单位定义为每分钟D值增加0.01, 单位为U·g-1·min-1.
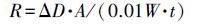
|
(1) |
式中, ΔD为反应时间内吸光度值的变化, A为稀释倍数, W为样品鲜重(g), t为反应时间(min).
3 结果和讨论(Results and discussion) 3.1 菲和芘的降解动力学菌株Pn2和033在30 ℃、pH=7.0条件下能够分别以菲和芘作为碳源和能源进行生长繁殖, 且对菲和芘具有高效的降解效能(图 2).接种0~1.5 d, 菌株Pn2的细胞数量(N)快速增加, 菲降解率也逐渐增加, 表明该菌能够以菲作为碳源和能源进行生长繁殖(图 2a);接种1.5 d后, 培养液中菌株Pn2的细菌数量呈现下降趋势, 菲降解率增加缓慢, 主要是因为降解液中菲含量较低, 菌株没有足够的碳源和能源进行生长繁殖;接种3 d后, 菌株Pn2对菲的降解率高达99.7%.菌株033对芘的降解动力学过程和规律与菌株Pn2类似, 接种6 d后, 其对芘的降解率高达98.3%(图 2b).依据培养液中PAHs残留浓度, 拟合降解动力学方程如下:C菲=134.49e-2.028t, 半衰期为0.34 d(R2=0.9889);C芘=57.226e-0.728t, 半衰期为0.95 d(R2=0.9861);其中, C表示培养基中PAHs残留浓度, t表示接种时间.
 |
| 图 2 PAHs降解菌的生长曲线及其对PAHs的降解动力学 (a.菲降解菌Pn2, b.芘降解菌033) Fig. 2 The biodegradation dynamics of PAHs and growth curve of single strain Pn2 or 033 with PAHs as their carbon source |
目前, 研究者已经从环境介质中筛选出多种具有PAHs降解特性的功能菌(徐成斌等, 2015; Bacosa et al., 2015; Liu et al., 2016;刁硕等, 2017).例如, 倪雪等(2013)从PAHs污染植物体内获得两株菲降解菌Stenotrophomonas sp. P1和Pseudomonas sp. P2, 其对菲的降解率均高达90%以上.李全霞等(2008)从土壤中分离一株高效芘降解菌Mycobacterium sp. M11, 该菌对菲、蒽和荧蒽等PAHs具有良好的降解效能.本研究中, 菌株Pn2和033分别对菲和芘具有高效的降解效率.已有资料显示, 功能菌株降解PAHs的过程主要包含2种关键酶, 一是起始反应阶段的双加氧酶, 它能够完成PAHs羟基化;二是邻苯二酚双加氧酶, 它可以催化单环芳烃氧化, 使苯环裂解并通过三羧酸循环矿化为CO2和H2O(Dean-Ross et al., 1996;王涛等, 2016;Waigi et al., 2017).菌株Pn2能够将菲的2个相邻碳原子羟基化形成顺-二醇, 并在多种酶系共同调节下导致苯环逐一裂解产生水杨酸和邻苯二酚(Liu et al., 2014);菌株033代谢芘的产物主要包括芘-4, 5-二醇、4, 5-二羧基-菲、4-菲甲酸和邻苯二甲酸(Dean-Ross et al., 1996).
3.2 接种混合菌降低黑麦草体内PAHs含量植物主要通过根部吸收POPs, 多数POPs的logKow值在3.0~8.3之间, 其值越低越容易被植物根吸收(Takaki et al., 2014).菲和芘的logKow值分别为4.57和5.18(图 1), 它们能够被黑麦草根高效地吸收积累, 并与根部脂质膜结合(Chaudhry et al., 2002).由于黑麦草体内缺少相应的PAHs转运蛋白, 其主要通过扩散作用将细胞壁中的菲和芘运输到木质部, 并由植物蒸腾拉力作用实现黑麦草根部PAHs向茎叶迁移(Campos et al., 2008).当黑麦草吸收转运PAHs时, 为避免PAHs对自身的毒害作用, 其能够通过蒸发或降解作用降低PAHs毒性(Arslan et al., 2017).本研究中, 接种混合菌株Pn2和033(CRB)可以作为一种新型有效途径用于增强黑麦草对PAHs的耐性, 甚至降低黑麦草体内PAHs污染.
如表 2所示, 接种混合PAHs降解菌株能够有效地减少黑麦草体内菲和芘的含量.与CRD(污染土壤+黑麦草+灭活的混合菌)处理组对比, 接种混合菌株Pn2和033促使黑麦草根和茎叶中菲含量分别由59.72和10.50 mg·kg-1降低至48.92和6.65 mg·kg-1, 芘含量分别由114.43和12.96 mg·kg-1降低至88.41和9.73 mg·kg-1(p < 0.05).黑麦草是畜牧业中常见牧草, 降低黑麦草体内PAHs污染对促进畜牧业发展、阻隔PAHs在植物-动物的食物链中传递及保护人群健康等具有重要意义(Sun et al., 2014a; Zhu et al., 2014).本研究中, 与CRD处理组对比, 接种混合菌株Pn2和033显著地降低了黑麦草体内21.63%的总PAHs含量.
| 表 2 在黄棕壤中种植黑麦草并接菌30 d后黑麦草体内菲和芘的含量 Table 2 The concentration of phenanthrene and pyrene in ryegrass planted in yellow-brown soil for 30 d |
黑麦草生物量的增加可能导致其体内PAHs含量减少, 计算黑麦草体内PAHs积累量可以排除该因素干扰(Sun et al., 2014b).黑麦草体内PAHs积累量(B)计算公式见式(2).
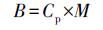
|
(2) |
式中, Cp表示植物体内PAHs含量(mg·kg-1), M表示植物干重(mg·盆-1).
由图 3可知, 与CRD处理组对比, 接种混合菌株Pn2和033(CRB)促使黑麦草根中菲和芘的积累量分别由1.69和4.18 μg·盆-1减少至1.34和2.37 μg·盆-1, 黑麦草茎叶中菲和芘的积累量分别由1.95和3.79 μg·盆-1减少至1.58和2.13 μg·盆-1.该研究结果进一步表明, 接种混合菌株Pn2和033能够降低黑麦草体内PAHs的含量.
 |
| 图 3 黑麦草根(a)和茎叶(b)中菲和芘的积累量 Fig. 3 The accumulation amounts of phenanthrene and pyrene in ryegrass roots (a) and shoots (b) |
转移系数(TF)的大小能够说明PAHs从黑麦草根向茎叶的传输能力, 进而反映植物地上部位PAHs污染风险(Sun et al., 2014b).TF计算公式如下所示:
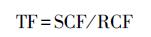
|
(3) |
式中, SCF表示PAHs在植物茎叶中的富集系数(SCF=Cs/Csoil), RCF表示PAHs在植物根中的富集系数(RCF=Cr/Csoil), Cs表示植物茎叶中PAHs含量(mg·kg-1), Cr表示植物根中PAHs含量(mg·kg-1), Csoil表示土壤中PAHs含量(mg·kg-1).
由图 4可知, 与CRD处理组对比, 接种混合菌株Pn2和033(CRB)促使黑麦草体内菲和芘的TF分别由0.1692和0.1216降低至0.1388和0.1191.这些结果表明, 接种混合菌株Pn2和033不仅能够降低黑麦草体内PAHs含量和积累量, 也可以降低PAHs由黑麦草根向茎叶的转移作用, 从而降低黑麦草地上部位PAHs污染风险.
 |
| 图 4 菲和芘在黑麦草体内的转移系数(TF) Fig. 4 The translocation factor (TF) of phenanthrene and pyrene in ryegrass |
接种混合功能菌株Pn2和033不仅能够降低植物体内PAHs污染, 也可以促进土壤中PAHs降解.与污染土壤接种灭活的混合菌相比(CD), CB(污染土壤+混合菌)、CRD和CRB(污染土壤+黑麦草+混合菌)均能够减少土壤中PAHs含量, 其中, CRB处理效果最佳, 致使土壤中菲和芘的去除率分别增加了37.3%和59.8%(图 5).研究人员采用PCR-DGGE技术证实, 不同浓度PAHs污染土壤中Massilia spp.的数量在微生物群落中始终占据主导地位, 其能够有效地降解土壤中PAHs(Zhang et al., 2010).Sun等(2014b)研究指出, 采用灌根法接种植物内生菌株Staphylococcus sp. BJ06能够促进黑麦草生长, 并有效地降低土壤-植物体系中芘含量.这些究结果表明, 植物-混合功能菌的联合作用能够高效地去除土壤中混合PAHs污染.土壤中PAHs含量的降低直接减少了植物对土壤中PAHs的吸收和积累, 导致植物体内PAHs含量下降(Sun et al., 2014b; Wang et al., 2017).
 |
| 图 5 不同处理土壤中菲和芘的含量 Fig. 5 The concentrations of phenanthrene and pyrene in soil by different treatments |
另有研究表明, 接种的外来功能菌株可能与土著微生物竞争或协同降解土壤中PAHs(Afzal et al., 2014).PAHs在植物体内的含量与土壤中PAHs含量呈正相关, 接种混合功能菌株Pn2和033可能在黑麦草根际土壤释放分泌液, 促进土壤中PAHs降解;同时, 两株菌的PAHs降解基因在土著微生物群落间的水平迁移也可以加速土壤中PAHs降解, 从而减少黑麦草对土壤中PAHs的吸收和积累(Taghavi et al., 2005).此外, 黑麦草根系产生的有机酸、根际氧化、共代谢诱导和生物表面活性剂等物质及功能, 不仅可以为混合功能菌株提供良好的生长环境, 也能够促进根际土壤中PAHs降解和矿化(Afzal et al., 2014; Sprocati et al., 2014).
3.4 接种混合菌影响植物体内PPO和POD活性植物体内POPs的降解主要通过氧化反应、水解作用和环氧化物的形成等途径实现(Chaudhry et al., 2002), 而这些过程需要POPs降解酶参与.研究指出, 植物体内的氧化酶、还原酶和酯酶等可以直接参与植物体内POPs的降解代谢过程(Kvesitadze et al., 2009);而接种功能菌可以通过诱导这些酶活性, 影响植物体内POPs代谢转移(Sinsabaugh, 2010;Afzal et al., 2014).如图 6所示, 所有种植黑麦草的处理组, 植物根中PPO和POD活性均高于茎叶.与CRD处理组对比, 接种混合菌株Pn2和033能够影响黑麦草体内PPO(p > 0.05)和POD(p < 0.05)活性.PAHs污染降低了黑麦草根中PPO活性, 可能是由于黑麦草根中PAHs含量过高, 对PPO活性起到抑制作用;而接种混合菌株显著地促进了黑麦草根和茎叶中POD活性, 其中, 根中POD活性最大, 为1611.62 U·g-1·min-1.这些结果表明, 当黑麦草受到PAHs污染胁迫时, 其体内PPO和POD活性发生改变, 从而介导植物对PAHs的应答反应和代谢作用;而接种混合功能菌株能够改变植物体内PAHs降解酶PPO和POD活性, 影响植物体内PAHs的降解代谢.
 |
| 图 6 菲和芘污染条件下黑麦草体内PPO(a)和POD酶活性(b) Fig. 6 The activities of PPO (a) and POD (b) in ryegrass under phenanthrene- and pyrene-contaminated conditions |
Weyens等(2009)研究指出, 接种菌株Burkholderia cepacia VM1468能够降低三氯乙烯污染的杨树体内接触酶和超氧化物歧化酶(SOD)活性.另有研究表明, 某些功能菌也能产生加氧酶作用于PAHs等有机污染物, 促使苯环裂解代谢(雷东锋等, 2004).因此, 通过接种混合功能菌群改变植物体内PAHs降解酶活性, 有望促进植物体内PAHs降解代谢.本研究中, 接种混合菌株Pn2和033可能通过诱导植物体内PAHs降解酶PPO和POD活性, 促进植物体内PAHs降解代谢.分析原因可能是由于POD活性的升高促进了植物体内超氧自由基的清除, 并保护植物细胞膜免受PAHs损伤、能够正常代谢, 进而直接增强植物对PAHs的利用效率(Kvesitadze et al., 2009;夏红霞等, 2013;Afzal et al., 2014).尽管如此, 今后的研究应当深入挖掘接种混合功能菌株对植物体内PAHs降解和相关酶活性变化的内在关系, 系统地揭示接种功能菌促进植物体内PAHs降解代谢的作用机理.
3.5 接种混合功能菌降低植物PAHs污染的作用机制结合本研究结果和已有的文献报道, 总结混合功能菌降低植物PAHs污染的作用机制主要包括以下5点:①接种混合功能菌直接降低了土壤中PAHs含量, 导致植物对土壤中PAHs的吸收和积累量降低(图 5);②接种混合功能菌显著地影响植物体内PAHs降解酶POD活性, 强化植物-混合功能菌对PAHs的降解代谢潜能(图 6);③混合功能菌定殖在植物细胞间隙和维管束导管中, 直接利用植物体内PAHs作为碳源和能源进行生长繁殖, 或以共代谢方式降解植物体内PAHs(Afzal et al., 2014; Sun et al., 2014a);④在植物根际或体内, 混合功能菌可以将PAHs降解基因水平迁移到其他土著菌群中, 促进整个根际或内生菌群对PAHs的降解代谢作用(Wilson et al., 2003; Wang et al., 2007);⑤混合功能菌PAHs降解基因在植物根际或体内的大量表达, 增加了PAHs降解基因的丰度和表达量, 促进了植物体内PAHs降解代谢潜能(Siciliano et al., 2001).
4 结论(Conclusions)1) 在30 ℃、pH=7.0条件下, 接种菌株Pn2和033对菲和芘的降解效率分别高达99.7%和98.3%, 降解半衰期分别为0.34 d和0.95 d.
2) 与CRD处理组对比, 接种混合菌株Pn2和033显著地降低了黑麦草体内PAHs含量, 并阻控黑麦草地上部位PAHs污染风险.
3) 接种混合菌株Pn2和033可增强黑麦草体内POD活性, 可能影响黑麦草体内PAHs的降解代谢.
Arslan M, Imran A, Khan Q M, et al. 2017. Plant-bacteria partnerships for the remediation of persistent organic pollutants[J]. Environmental Science and Pollution Research, 24(5): 4322–4336.
DOI:10.1007/s11356-015-4935-3
|
Afzal M, Khan Q M, Sessitsch A. 2014. Endophytic bacteria:Prospects and applications for the phytoremediation of organic pollutants[J]. Chemosphere, 117: 232–242.
DOI:10.1016/j.chemosphere.2014.06.078
|
Bacosa H P, Inoue C. 2015. Polycyclic aromatic hydrocarbons (PAHs) biodegradation potential and diversity of microbial consortia enriched from tsunami sediments in Miyagi, Japan[J]. Journal of Hazardous Materials, 283: 689–697.
DOI:10.1016/j.jhazmat.2014.09.068
|
Barnier C, Ouvrard S, Robin C, et al. 2014. Desorption kinetics of PAHs from aged industrial soils for availability assessment[J]. Science of the Total Environment, 470: 639–645.
|
Çakmakçi R, Dönmez F, Aydın A, et al. 2006. Growth promotion of plants by plant growth-promoting rhizobacteria under greenhouse and two different field soil conditions[J]. Soil Biology & Biochemistry, 38(6): 1482–1487.
|
Cai C Y, Li J Y, Wu D, et al. 2017. Spatial distribution, emission source and health risk of parent PAHs and derivatives in surface soils from the Yangtze River Delta, eastern China[J]. Chemosphere, 178: 301–308.
DOI:10.1016/j.chemosphere.2017.03.057
|
Campos V, Merino I, Casado R, et al. 2008. Phytoremediation of organic pollutants[J]. Spanish Journal of Agricultural Research, 6: 38–47.
DOI:10.5424/sjar/200806S1-372
|
Chen Z X, Ni H G, Jing X, et al. 2015. Plant uptake, translocation, and return of polycyclic aromatic hydrocarbons via fine root branch orders in a subtropical forest ecosystem[J]. Chemosphere, 131: 192–200.
DOI:10.1016/j.chemosphere.2015.03.045
|
Chaudhry Q, Schröder P, Werck-Reichhart D, et al. 2002. Prospects and limitations of phytoremediation for the removal of persistent pesticides in the environment[J]. Environmental Science and Pollution Research, 9: 4–17.
DOI:10.1007/BF02987313
|
陈保冬, 赵方杰, 张莘, 等. 2015. 土壤生物与土壤污染研究前沿与展望[J]. 生态学报, 2015, 35(20): 6604–6613.
|
刁硕, 王红旗, 许洁, 等. 2017. 低温耐盐芘降解菌的筛选鉴定及降解特性研究[J]. 中国环境科学, 2017, 37(2): 677–685.
|
Dean-Ross D, Cerniglia C E. 1996. Degradation of pyrene by Mycobacterium flavescens[J]. Applied Microbiology and Biotechnology, 46: 307–312.
DOI:10.1007/s002530050822
|
Glick B R. 2014. Bacteria with ACC deaminase can promote plant growth and help to feed the world[J]. Microbiological Research, 169(1): 30–39.
DOI:10.1016/j.micres.2013.09.009
|
Gao Y, Zhu L. 2004. Plant uptake, accumulation and translocation of phenanthrene and pyrene in soils[J]. Chemosphere, 55(9): 1169–1178.
DOI:10.1016/j.chemosphere.2004.01.037
|
Iwabuchi T, Harayama S. 1998. Biochemical and molecular characterization of 1-hydroxy-2-naphthoate dioxygenase from Nocardioides sp.KP7[J]. Journal of Biological Chemistry, 273(14): 8332–8336.
DOI:10.1074/jbc.273.14.8332
|
Kim K H, Jahan S A, Kabir E, et al. 2013. A review of airborne polycyclic aromatic hydrocarbons (PAHs) and their human health effects[J]. Environment International, 60: 71–80.
DOI:10.1016/j.envint.2013.07.019
|
Kvesitadze E, Sadunishvili T, Kvesitadze G. 2009. Mechanisms of organic contaminants uptake and degradation in plants[J]. World Academy of Science, Engineering and Technology, 55(6): 458–468.
|
Liu J, Liu S, Sun K, et al. 2014. Colonization on root surface by a phenanthrene-degrading endophytic bacterium and its application for reducing plant phenanthrene contamination[J]. PloS One, 9(9): e108249.
DOI:10.1371/journal.pone.0108249
|
Liu J, Xiang Y, Zhang Z, et al. 2017. Inoculation of a phenanthrene-degrading endophytic bacterium reduces the phenanthrene level and alters the bacterial community structure in wheat[J]. Applied Microbiology and Biotechnology, 101(12): 5199–5212.
DOI:10.1007/s00253-017-8247-z
|
Liu S H, Zeng G M, Niu Q Y, et al. 2016. Bioremediation mechanisms of combined pollution of PAHs and heavy metals by bacteria and fungi:A mini review[J]. Bioresource Technology, 224: 35–33.
|
雷东锋, 冯怡, 蒋大宗. 2004. 植物中多酚氧化酶的特征[J]. 自然科学进展, 2004, 14(6): 606–614.
DOI:10.3321/j.issn:1002-008X.2004.06.002 |
李全霞, 范丙全, 龚明波, 等. 2008. 降解芘的分枝杆菌M11的分离鉴定和降解特性[J]. 环境科学, 2008, 29(3): 763–768.
DOI:10.3321/j.issn:0250-3301.2008.03.037 |
刘爽, 刘娟, 凌婉婷, 等. 2013. 一株高效降解菲的植物内生细菌筛选及其生长特性[J]. 中国环境科学, 2013, 33(1): 95–102.
DOI:10.3969/j.issn.1000-6923.2013.01.014 |
Menzie C A, Potocki B B, Santodonato J. 1992. Exposure to carcinogenic PAHs in the environment[J]. Environmental Science & Technology, 26: 1278–1284.
|
倪雪, 刘娟, 高彦征, 等. 2013. 2株降解菲的植物内生细菌筛选及其降解特性[J]. 环境科学, 2013, 34(2): 746–752.
|
Sprocati A, Alisi C, PintoV, et al. 2014. Assessment of the applicability of a "toolbox" designed for microbially assisted phytoremediation:The case study at Ingurtosu mining site (Italy)[J]. Environmental Science and Pollution Research, 21: 6939–6951.
DOI:10.1007/s11356-013-2154-3
|
Sinsabaugh R L. 2010. Phenol oxidase, peroxidase and organic matter dynamics of soil[J]. Soil Biology and Biochemistry, 42(3): 391–404.
DOI:10.1016/j.soilbio.2009.10.014
|
Siciliano S D, Fortin N, Mihoc A, et al. 2001. Selection of specific endophytic bacterial genotypes by plants in response to soil contamination[J]. Applied and Environmental Microbiology, 67(6): 2469–2475.
DOI:10.1128/AEM.67.6.2469-2475.2001
|
Shah K, Kumar R G, Verma S, et al. 2001. Effect of cadmium on lipid peroxidation, superoxide anion generation and activities of antioxidant enzymes in growing rice seedlings[J]. Plant Science, 161(6): 1135–1144.
DOI:10.1016/S0168-9452(01)00517-9
|
Sun K, Liu J, Gao Y, et al. 2014a. Isolation, plant colonization potential, and phenanthrene degradation performance of the endophytic bacterium Pseudomonas sp.Ph6-gfp[J]. Scientific Reports, 4: 5462.
|
Sun K, Liu J, Jin L, et al. 2014b. Utilizing pyrene-degrading endophytic bacteria to reduce the risk of plant pyrene contamination[J]. Plant and Soil, 374(1/2): 251–262.
|
Samsøe-Petersen L, Larsen E H, Larsen P B, et al. 2002. Uptake of trace elements and PAHs by fruit and vegetables from contaminated soils[J]. Environmental Science & Technology, 36(14): 3057–3063.
|
盛月慧, 刘娟, 高彦征, 等. 2013. 黑麦草体内POD和PPO活性及可培养内生细菌种群对不同浓度菲污染的响应[J]. 南京农业大学学报, 2013, 36(6): 51–59.
|
孙凯, 刘娟, 李欣, 等. 2014. 两株具有芘降解功能的植物内生细菌的分离筛选及其特性[J]. 生态学报, 2014, 34(4): 853–861.
|
Taghavi S, Barac T, Greenberg B, et al. 2005. Horizontal gene transfer to endogenous endophytic bacteria from poplar improves phytoremediation of toluene[J]. Applied and Environmental Microbiology, 71(12): 8500–8505.
DOI:10.1128/AEM.71.12.8500-8505.2005
|
Takaki K, Wade A J, Collins C D. 2014. Assessment of plant uptake models used in exposure assessment tools for soils contaminated with organic pollutants[J]. Environmental Science & Technology, 48: 12073–12082.
|
Wilson M S, Herrick J B, Jeon C O, et al. 2003. Horizontal transfer of phnAc dioxygenase genes within one of two phenotypically and genotypically distinctive naphthalene-degrading guilds from adjacent soil environments[J]. Applied and Environmental Microbiology, 69(4): 2172–2181.
DOI:10.1128/AEM.69.4.2172-2181.2003
|
Wang J, Liu J, Ling W, et al. 2017. Composite of PAH-degrading endophytic bacteria reduces contamination and health risks caused by PAHs in vegetables[J]. Science of the Total Environment, 598: 471–478.
DOI:10.1016/j.scitotenv.2017.04.126
|
Waigi M G, Sun K, Gao Y. 2017. Sphingomonads in microbe-assisted phytoremediation:Tackling soil pollution[J]. Trends in Biotechnology, 35(9): 883–899.
DOI:10.1016/j.tibtech.2017.06.014
|
Wang Y, Xiao M, Geng X, et al. 2007. Horizontal transfer of genetic determinants for degradation of phenol between the bacteria living in plant and its rhizosphere[J]. Applied Microbiology and Biotechnology, 77(3): 733–739.
DOI:10.1007/s00253-007-1187-2
|
Wang Y, Tian Z, Zhu H, et al. 2012. Polycyclic aromatic hydrocarbons (PAHs) in soils and vegetation near an e-waste recycling site in South China:Concentration, distribution, source, and risk assessment[J]. Science of the Total Environment, 439: 187–193.
DOI:10.1016/j.scitotenv.2012.08.018
|
Weyens N, Van Der Lelie D, Artois T, et al. 2009. Bioaugmentation with engineered endophytic bacteria improves contaminant fate in phytoremediation[J]. Environmental Science & Technology, 43(24): 9413–9418.
|
王涛, 蓝慧, 田云, 等. 2016. 多环芳烃的微生物降解机制研究进展[J]. 化学与生物工程, 2016, 33(2): 8–14.
DOI:10.3969/j.issn.1672-5425.2016.02.002 |
夏红霞, 朱启红. 2013. Pb、Zn及其交互作用对蜈蚣草抗氧化酶及叶绿素含量的影响[J]. 环境化学, 2013, 32(6): 959–963.
|
徐成斌, 王闻烨, 李鲜珠, 等. 2015. 一株菲降解菌的鉴定及降解特性[J]. 环境科学学报, 2015, 35(3): 684–691.
|
Zhu X, Ni X, Liu J, et al. 2014. Application of endophytic bacteria to reduce persistent organic pollutants contamination in plants[J]. CLEAN-Soil Air Water, 42(3): 306–310.
DOI:10.1002/clen.v42.3
|
Zhu X, Ni X, Waigi M G, et al. 2016. Biodegradation of mixed PAHs by PAH-degrading endophytic bacteria[J]. International Journal of Environmental Research and Public Health, 13(8): 805.
DOI:10.3390/ijerph13080805
|
Zielińska A, Oleszczuk P. 2015. Evaluation of sewage sludge and slow pyrolyzed sewage sludge-derived biochar for adsorption of phenanthrene and pyrene[J]. Bioresource Technology, 192: 618–626.
DOI:10.1016/j.biortech.2015.06.032
|
Zhang W, Wang H, Zhang R, et al. 2010. Bacterial communities in PAH contaminated soils at an electronic-waste processing center in China[J]. Ecotoxicology, 19: 96–104.
DOI:10.1007/s10646-009-0393-3
|