 2019, Vol. 39
2019, Vol. 39



2. 福建师范大学地理研究所, 福州 350007;
3. 福建师范大学地理科学学院, 福州 350007;
4. 福建师范大学亚热带湿地研究中心, 福州 350007;
5. 福州大学环境与资源学院, 福州 350007;
6. 国家林业和草原局福建闽江河口湿地生态系统定位观测研究站, 福州 350007
2. Institute of Geography, Fujian Normal University, Fuzhou 350007;
3. School of Geographical Sciences, Fujian Normal University, Fuzhou 350007;
4. Research Centre of Wetlands in Subtropical Region, Fujian Normal University, Fuzhou 350007;
5. College of Environment and Resources, Fuzhou University, Fuzhou 350007;
6. Wetland Ecosystem Research Station of Minjiang River Estuary, State Administration of Forestry and Grassland, Fuzhou 350007
红树林凋落物产量和分解动态变化对亚热带海岸生态系统物质循环和能量转换具有重要贡献(Silva et al, 1998; Sánchez-Andrés et al, 2010).凋落物的分解和营养物质的高效循环又为红树林生态系统的高生产力提供了保证(Bosire et al, 2005).由此, 凋落物产量、养分含量特征及分解动态成为评估红树林生态系统生产力的重要参考依据(Ye et al, 2013; Numbere et al., 2017).
研究发现, 凋落物是红树林生态系统的主要营养来源(Tam et al, 1998), 不同研究区域的红树林树种、土壤养分状况和环境因子都对红树林凋落物的产量有重要影响(Jr et al, 1996;Silva et al, 1998;Feller et al, 1999; Arreola-Lizárraga et al, 2004).中国亚热带红树林一年内凋落物产量高峰期一般出现在多雨的夏季(Tam et al., 1998).碳(C)、氮(N)和磷(P)是维持植物生长发育所必需的营养元素(Ågren, 2008), 植物从土壤中获取营养, 又以凋落物的形式将养分归还给土壤, 实现生态系统养分循环过程(Yang et al, 2014).因此, 研究凋落物养分含量及其分解动态对红树林生态系统的物质循环有着重要意义.
凋落物的分解是凋落物经过物理破碎和化学过程最终转化为碳水化合物并释放出能量的过程(Chapman, 1976), 而凋落物分解速率受气温、降水、凋落物质量及性质的影响显著(Li et al, 2011; Salah et al., 2011).研究表明, 随着分解不断进行, 凋落物质量发生变化并影响到分解速率和元素的释放(Attiwill et al., 1993);凋落物元素的含量, 尤其是C、N、P元素及其化学计量比对凋落物的分解起到重要的指示作用(曲浩等, 2010).同时, 凋落物的分解还受到凋落物酶活性的影响, 因此,凋落物的分解是在凋落物与酶系统的相互作用下完成的(葛晓改等, 2014).例如, 凋落物分解过程中氮的输入有利于提高纤维素水解酶、β-葡萄糖苷酶等水解酶的活性, 从而加速凋落物的分解(Sinsabaugh et al, 2002);且有研究表明, 凋落物的分解过程需要多种酶的共同参与(Waring, 2013;季晓燕等, 2013a).凋落物分解与酶活性相互作用机制是凋落物分解研究的热点问题(Zhang et al., 2006), 但目前国内外有关红树林凋落物分解过程中酶活性的相关报道较少(Wang et al, 2013).
红树林主要位于热带和亚热带海岸地带, 凋落叶分解受季节变化和潮汐作用的影响较大(Twilley, 1985; Wafar et al, 1997; Dick et al, 2000; Bosire et al, 2005).目前, 国外有关红树林凋落物及其分解已有大量报道, 主要针对美国加利福尼亚州、巴西、印度和莫桑比克共和国等地(Silva et al, 1998; Fernando et al., 2009; Sánchez-Andrés et al, 2010; Chanda et al, 2015), 我国针对福建、广东、广西和海南等地典型红树林树种凋落物产量动态也开展了大量工作, 为红树林生态系统凋落物量的研究提供了丰富的参考性数据(郑逢中等, 1998;张乔民等, 2003;朱可峰等, 2011).红树林具有良好的促淤作用, 有的凋落物在被沉积物掩埋后进行分解, 由于地表温度较高, 蒸发量大使得凋落物持水量少, 而地下温度一般低于地表, 沉积物覆盖使凋落物保持湿润状态, 有利于微生物对其进行分解(Garciapausas et al, 2004).盛华夏(2009)在研究九龙江口秋茄凋落叶滩面分解(地上组)和埋于沉积物5 cm处(地下组)分解的动态时发现, 两个处理的分解动态存在差异, 但并未达到显著性水平.对珠海淇澳人工红树林凋落物分解的研究也发现, 凋落物分解与气温、土温关系密切(杨秀, 2015).
尽管国外对红树林凋落物产量及其分解的研究已有较多报道(Fernando et al., 2009; Sánchez-Andrés et al, 2010; Silva et al, 2007), 但有关红树林凋落物分解过程中水解酶活性的研究较为少见, 而凋落物水解酶活性对凋落物的分解有重要影响(Alarcón-Gutiérrez et al., 2010;季晓燕等, 2013a).因此, 本文在前人研究的基础上, 以福建省闽江口20年生人工种植秋茄林(Kandelia obovata)为研究对象, 研究秋茄凋落物年产量及其养分含量动态和凋落叶分解元素动态及其影响因素, 其中, 凋落物分解设置平铺于红树林下沉积物表面(简称:地上组AS)和埋于沉积物以下5 cm深度处(简称:地下组BS)两个处理, 模拟自然状态下沉积作用对凋落物分解的影响, 对比分析分解过程中元素的动态特征及土壤环境因子对凋落物酶活性的影响, 以期为亚热带河口红树林生态系统碳、氮、磷元素循环和养分平衡提供参考.
2 研究方法(Research methods) 2.1 研究区概况研究区位于福建省闽江口粗芦岛(119°38′02″E, 26°08′27″N), 濒临东海, 平均海拔4 m, 属亚热带海洋性季风气候(图 1), 年均气温19.7 ℃, 年均降水量1346 mm.粗芦岛分布着河口半咸水湿地, 主要植被类型包括土著种芦苇(Phragmites australis (Cav.) Trin. ex Steud)、外来入侵种互花米草(Spartina alterniflora Loisel.)和20年生人工种植秋茄林.秋茄红树林与岸线平行呈带状分布, 平均株高为(3.86±0.05) m, 胸径为(3.69±0.12) cm, 冠幅为(0.9±0.1) m, 密度为23500株·hm-2.
 |
| 图 1 研究区及采样点布设示意图 Fig. 1 Location of study area and sampling site |
2017年1—12月, 在秋茄林内共布设6个凋落物收集框, 为避免涨潮淹没收集框, 设置每个收集框出露地面约2 m高度, 每两个收集框之间需保持10 m间距并随机分布于秋茄林内(图 1).收集框由铝合金材料制成, 缝上尼龙网, 孔径为1.5 mm, 网口面积为0.6 m×0.6 m, 深约30 cm, 每隔一个月收集1次凋落物, 共72个网次.
2.2.2 凋落物分解实验设计与采样在实验室, 选取黄熟凋落叶用蒸馏水洗净擦干, 使用万分之一天平称取凋落叶约20 g(鲜重), 并记录称取质量的精确数值, 然后平铺于20 cm×20 cm的尼龙网分解袋(1 mm×1 mm孔径)并封好(Numbere et al., 2017).在秋茄样地内设置3个凋落物分解袋投放点, 每两个采样点依次间距10 m(图 1), 用PVC管作固定桩, 用尼龙绳将分解袋系于固定桩.
2017年3月30日开始投放分解袋, 共准备87个分解袋, 随机选取3袋不予投放, 用于初始质量的测定.在3个凋落物投放点中, 每个投放点投放28个分解袋, 实验分为两个处理, 其中, 地上组(Above the soil surface):投放14个(2个备用), 分散平铺于林地表面, 简称AS;地下组(Below the soil surface):投放14个(2个备用), 分散埋藏于林地表面以下5 cm深处, 简称BS.样地内共投放84个分解袋(其中12个备用), 投放后的第7、16、29、60、90、120 d分6次分别取回12个分解袋.每个投放点地上、地下分别采集2袋, 分别用于测定凋落物质量、碳、氮、磷含量和凋落物水解酶活性.采样时使用2265FS便携式电导计(Spectrum Technologies Inc, USA)测定每个采样点的土壤温度和电导率.同时,在样地内设置一个百叶箱,箱内装有温湿度记录仪(RC-4温度记录仪),每隔2 h记录一次秋茄林内气温和相对湿度(图 2).
 |
| 图 2 2017年1月1日—12月30日研究样地的气温和相对湿度 Fig. 2 Dynamics of air temperature and humidity at the location of study sampling site from January 1, 2017 to December 30, 2017 |
将每月收集到的凋落物挑拣出鸟类羽毛、粪便等杂物后, 用干毛刷清除凋落物表面灰尘, 按叶、枝、花和果实(包括胚轴)4个组分分开, 于75 ℃烘箱烘干至恒重, 记录当月凋落物各组分产量和总产量.
2.3.2 凋落物分解样品处理与测定凋落物质量测定时需将样品洗净擦干称取鲜重, 于75 ℃烘箱烘至恒重后记录干重;之后研磨过100目网筛, 用碳氮元素分析仪(Vario EL Ⅲ, 德国)测定凋落物TC和TN;凋落物TP测定采用HClO4-H2SO4消煮法, 并用连续流动分析仪(SKALAR San++, Netherlands)测定;用于凋落物酶活性测定的样品于4 ℃冰箱保存, 将新鲜凋落物剪碎为约1 cm2的碎片, 使用万分之一天平称取每个样品约0.5 g并记录准确数值, 之后加入125 mL醋酸缓冲液(浓度为5 mmol·L-1, pH=5), 用磁力搅拌器搅拌约5 min使其均匀混合.用移液枪取上清液200 μL于96微孔板, 加入反应所需底物:β-葡萄糖苷酶(β-1, 4-glucosidase)、纤维素水解酶(Cellobiohydrolase)、β-N-乙酰氨基葡萄糖苷酶(β-1, 4-N-acetylglucosaminidase)和酸性磷酸酶(Acid phosphatase), 4种水解酶(以下分别简称βG、CBH、NAG和AP)的活性底物分别为4-MUB-β-D-glucoside、4-MUB-β-D- cellobioside、4-MUB-N-acetyl-β-D-glucosaminide和4-MUB-phosphate, 将样品于20 ℃避光培养4 h后向每个微孔加入10 μL的氢氧化钠溶液(浓度为1.0 mol·L-1)使反应停止, 每次测量样品均有16个重复.使用多功能酶标仪(SynergyH4, 美国)测定酶活性, 采用365 nm激发光和450 nm发射光滤光器检测荧光度, 阴性对照和淬火标准液校正后, 酶活性以每小时每克干物质产生底物的物质的量(nmol·h-1·g-1)表示(Saiya-Cork et al., 2002, Sinsabaugh et al., 2005).
2.4 数据处理与分析凋落叶分解过程中干物质残留率计算公式如下:
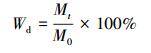
|
(1) |
式中, Wd为t时刻凋落叶干物质残留率, Mt为分解t时刻剩余干物质质量(g), M0为凋落叶初始时刻干物质质量(g).
使用Excel 2013对原始数据进行处理.使用SPSS 19.0中的单因素方差分析(One-way ANOVA)的最小显著差异法(LSD)对凋落物C、N、P含量月动态变化进行比较, 利用Paired-Samples T检验比较不同处理之间凋落叶干重残留率及分解速率的差异, 并采用一元线性回归分析建立凋落物残留率的自然对数与分解天数的回归方程, 采用重复测量方差分析对凋落物残留率、元素动态特征和水解酶活性进行分析, 凋落物元素含量和土壤环境因子与水解酶活性的相关性分析采用Pearson相关性分析, 采用多元线性逐步回归法分析凋落物元素含量和土壤环境因子对水解酶活性的影响并建立最优方程.使用Origin 8.5进行相关绘图.文中数据均表示为平均值±标准误差(mean±SE).
3 结果与分析(Results and analysis) 3.1 凋落物产量秋茄凋落物年产量为618.79 g·m-2·a-1, 各组分占总产量的比例分别为61.2%(叶)、10.5%(枝)、4.6%(花)、23.7%(果).由图 3可以看出, 凋落物产量具有明显的季节变化规律, 凋落物产量的峰值出现在8月, 5月次之, 11月的凋落物产量最少.
 |
| 图 3 2017年1—12月凋落物各组分的月变化 Fig. 3 Monthly dynamics of the different components of litter fall from January to December, 2017 |
落叶在凋落物中占据主体, 其变化趋势与凋落物总产量变化一致.落叶、落枝和落花量8月最多, 落果量5月最多, 其中4月出现落叶量的次峰值.除7、8月外, 落枝量在其它月份较少且趋于平缓.落花和落果在非生产期产量为零.
3.2 凋落物叶片C、N、P含量及其化学计量比由图 4可知, 凋落物叶片TC含量平均为(471.0±3.2) g·kg-1, 以12月最高, 3月最低;TN含量在8月((15.1±2.1) g·kg-1)显著高于其它月份(p < 0.05), 全年平均含量为(9.6±0.5) g·kg-1;TP含量在1—3月显著高于其他月份, 全年平均含量为(0.5±0.1) g·kg-1;1月C/N值最高, 为60.8, 8月C/N值(31.2)显著低于其他月份(p < 0.05);N/P和C/P在9月显著升高(p < 0.05), 分别为111.8和5529.0, 12月有一个次峰.
 |
| 图 4 凋落叶C、N、P含量及其化学计量比动态特征(n=3;不同小写字母表示不同月份同一指标差异显著) Fig. 4 Dynamics of litter fall C, N, P contents and their stoichmetric ratios |
由图 5可知, 随着分解的进行, 地上组和地下组两处理之间干物质残留率差异不显著(p>0.05).重复测量方差分析结果表明, 秋茄凋落叶干物质残留率随着分解时间的增加而显著降低(p < 0.01), 但在处理与时间的交互作用下未达到显著性水平(p>0.05).由凋落物残留率的自然对数与分解时间的回归分析结果(表 1)可知, 地上组(AS)和地下组(BS)半分解期分别为80.4 d和80.3 d.
 |
| 图 5 各处理之间干物质残留率 (n=3;TI:分解时间的影响;TR:处理的影响;TI×TR:时间和处理的交互作用影响;*和**分别表示在0.05和0.01水平上差异显著,下同) Fig. 5 Dry mass remaining of litter in treatment |
| 表 1 凋落物残留率(y)的自然对数与分解天数(x)的回归方程及相应参数 Table 1 Equations and parameters of the natural logarithm of litter mass remaining(y) regressed on decomposition days (x) above the soil surface(AS) and below the soil surface(BS) |
由图 6可知, TC含量呈先上升后下降的趋势, 在分解第29 d以后地上组含量大于地下组;TN含量呈不断上升的趋势, 整个分解期间地上组含量大于地下组;C/N呈现不断下降的趋势, 且地上组的比值小于地下组;TP含量呈先下降后升高的趋势, C/P和N/P呈先升高后下降的趋势.重复测量方差分析结果表明, 秋茄凋落叶C、N、P含量及其化学计量比随时间变化均表现出显著性差异(p < 0.01).其中, TC、TN、C/N和C/P在不同处理之间差异性显著(p < 0.05), 且时间和处理的交互作用对TC、TN、C/P和N/P的影响达到显著性水平(p < 0.05).
 |
| 图 6 凋落叶分解过程中碳、氮、磷含量及其化学计量比(n=3) Fig. 6 Dynamics of litterfall C, N, P contents and their stoichiometric ratios during litter decomposition |
由表 2可知, 地上组土壤电导率、温度与凋落叶TC含量、C/N、C/P呈负相关关系(p < 0.05, p < 0.01), 与TN含量呈正相关关系(p < 0.05, p < 0.01).地下组土壤温度与凋落叶TN含量呈正相关关系, 与C/N呈负相关关系(p < 0.05).地上组和地下组凋落叶N/P均与土壤电导率呈负相关关系(p < 0.05, p < 0.01).
| 表 2 凋落物元素含量与土壤环境因子的相关分析(n=15) Table 2 Pearson correlation between element content of litter and soil environmental factors |
由图 7可以看出, βG、CBH和NAG水解酶活性均呈现先升高后降低的趋势, 且均在分解的第60 d达到最大值.AP活性呈现波动上升的趋势, 在第120 d达到最高值.重复测量方差分析结果表明, 4种水解酶随时间变化均表现出显著性差异(p < 0.01), 且CBH在处理(TR)及时间×处理(TI×TR)的交互作用的影响下达到显著性水平(p < 0.05).
 |
| 图 7 凋落叶分解过程中水解酶活性动态变化(n=3) Fig. 7 Dynamics of enzyme activities during litter decomposition |
由表 3可知, 地上组和地下组AP水解酶活性均与凋落叶TC、C/N和C/P存在负相关关系(p < 0.05), 与TN、TP含量和土壤温度呈正相关关系(p < 0.05), 地上组AP水解酶活性与土壤电导率呈正相关关系(p < 0.05), CBH水解酶活性与C/P呈正相关关系(p < 0.05).利用多元逐步回归法分析凋落叶水解酶活性的主要影响因子, 得到水解酶活性与土壤温度、电导率、凋落叶碳、氮、磷含量及碳氮比之间的最优方程(表 4).结果表明, 土壤电导率对地上组AP水解酶活性影响较大, 土壤温度和凋落叶TP含量是影响地下组AP水解酶活性的主要因子.
| 表 3 水解酶活性与凋落叶元素动态和土壤环境因子的相关分析(n=15) Table 3 Pearson correlation between enzyme activities of litter and element content and soil environmental factors |
| 表 4 酸性磷酸酶活性与凋落叶元素动态和土壤环境因子的回归方程(n=15) Table 4 Multiple regression between activities of acid phosphatase and element content and soil environmental factors |
本研究中, 秋茄凋落物各组分产量随季节变化而不同, 8月总产量最高, 且当月落花、落枝和落叶的产量也达到峰值.秋茄落花期出现在7—9月, 对于福建省九龙江口秋茄凋落物产量的研究发现, 秋茄落花产量最高出现在7月, 这可能是由于研究期间当月风力大小和降雨量不同等天气状况造成的误差(Mfilinge et al, 2005).本研究秋茄果实产量出现在4—6月, 最高值出现在5月, 这与九龙江口秋茄凋落物产量动态研究结果一致, 且落花和落果的季节特征由不同物种特殊的生物特征决定(Ye et al., 2013).由于冬季气温较低使红树植物生长缓慢、生产力降低, 导致凋落物产量降低, 本研究凋落物产量最低值出现在11月, 这与以往的研究结果一致(Mackey et al., 1996; Mfilinge et al, 2005).国内外有关红树林凋落物年产量研究的范围在572.0~1640.82 g·m-2之间(Hoque et al, 2015), 相比其他森林凋落物年产量要高(邱明红等, 2017), 说明红树林生态系统具有高生产率和高归还率.本研究秋茄凋落物年产量为618.79 g·m-2·a-1, 与国内其他同类型红树植物凋落物年产量相差不大(郑逢中等, 1998).各组分中, 落叶占凋落物总量的比重最大, 这与国内外相关研究结果一致(Sánchez-Andrés et al, 2010;卢伟志等, 2014).落枝会受到风力影响, 比重次之, 而落花和落果受到植物生长规律的限制, 只在特定的月份出现, 其它月份产量多为零(Imgraben et al., 2008).本研究除落果占总质量的比重(23.7%)超过落枝(10.5%)与以往的研究不同外, 凋落物各组分量月动态变化与国内同纬度带秋茄凋落物量的研究结果一致(郑逢中等, 1998;金亮等, 2016).有研究认为森林凋落物量与高温和缺水密切相关, 植物必须通过自疏作用放弃大量的枝和叶来应对缺水状况, 或出现在多雨的季节, 雨水中携带了大量的营养物质使凋落物具有较高的生产量(Lee, 1989a;Wafar et al, 1997).由于红树林生长在盐度较高的沿海地带, 再加上夏季的高温, 蒸腾作用比较强烈, 这也是造成红树林凋落物产量较高的原因之一(Wium-Andersen, 1981), 台风等极端天气过后也会导致凋落物量有所增加.总之, 影响红树林凋落物量的因素复杂多样(张乔民等, 2003), 本研究收集了福建省闽江口秋茄凋落物, 这为了解亚热带红树林植物凋落物量和组分月动态变化提供了一定的参考数据, 但由于研究时间仅为一年, 存在一定的局限性, 因此, 只有保持长期监测才能清楚地认识其规律, 了解红树林的生态价值.
凋落物的C、N、P含量对于森林生态系统的元素循环有重要意义(姜沛沛等, 2016), 凋落物的养分含量随季节变化而不同(Bosire et al., 2005).本研究结果表明, 凋落叶TN含量8月显著高于其他月份, 而当月凋落叶产量达到一年中的最高峰值, 说明秋茄凋落叶在8月能够归还给土壤较高的含氮养分, 这与不同生长阶段的植物体养分的分配特征有关, 8月秋茄落叶、落花和落枝在一年中产量最高, 处于生长旺盛期, 各器官的养分含量相对其他月份较高(Güsewell, 2004).有研究表明, 衰老黄熟叶片的C/P和N/P随气温的增加而增加(Zheng et al., 2007), 这与本研究结论一致, 气温升高加快了植物体的新陈代谢速率, 使秋茄叶片所需养分含量升高, 含氮物质增加, 以维持生长代谢的需要.此外, 凋落物叶片养分含量的季节变化还受到树的种类、气候条件和所处地理位置的影响(Zheng et al., 2007).而国内外有关红树林树种凋落叶养分动态变化的相关性研究较为少见(Silva et al, 1998; Ye et al., 2013), 了解红树林凋落叶C、N、P养分含量的季节变化对认识红树林植物的自然节律特征和生态系统养分利用状况有着重要意义.同时, 通过本研究希望能为我国红树林凋落物养分的季节变化规律提供参考性数据.
4.2 凋落物分解动态由于长期周期性潮汐涨落, 红树林凋落物在分解前期具有较高的分解速率和淋溶速率(Valk et al., 1984; Mfilinge et al., 2005).本研究结果表明, 地上组(AS)和地下组(BS)半分解期分别为80.4 d和80.3 d, 且地上组与地下组两处理的凋落物分解干物质残留率无显著差异, 但二者随分解时间均表现出显著性差异.有研究表明, 白骨壤凋落叶干季和湿季半分解期分别为96 d和78 d(Fernando et al., 2009), 这与本研究结果中秋茄凋落叶半分解期时间接近.福建省漳江口秋茄凋落叶半分解期为50 d(Zhou et al., 2012), 也有研究发现福建省九龙江口秋茄凋落叶夏季和冬季的半分解期分别为18.7 d和47.4 d(Li et al., 2014), 而本研究秋茄凋落叶半分解期均大于以上研究, 这与研究区所处纬度、地理位置和采样季节有关, 不同纬度地区气候条件不同, 较高的温度条件下微生物活性较高, 有利于凋落物的分解(Mackey et al., 1996).本研究区所处纬度高于以上研究区, 年平均气温低于以上研究区, 因此, 凋落物表面微生物活性偏低, 分解时间较长.有研究表明, 初始N、P含量高的凋落叶会促进微生物对养分的吸收, 加快分解进程(Fenchel et al., 1999);研究发现, 较高树龄(24年生)的凋落物叶片养分含量往往低于较低树龄(11年生), 且较高树龄的凋落叶木质素含量较高不利于分解(Nga et al., 2005; Li et al., 2014).此外, 研究区所处潮滩高度及淹水频率也是影响凋落物分解的因素(Lee, 1989b; Mackey et al., 1996; Dick et al., 2000; Mfilinge et al., 2002).
湿地凋落物的分解分为“可溶性物质的淋溶”、“碎屑物质的微生物降解”和“物理和生物的分解”3个阶段, 这3个阶段与凋落物元素含量的变化密切相关(Valiela et al., 1985).本研究中在分解的第29 d以后地上组TC含量显著小于地下组(p < 0.05), 这可能与地上组因频繁被潮汐冲刷, 可溶性有机物不断被淋溶及凋落物碎屑的流失有关(张川等, 2016).随着凋落物分解的进行, 地上组和地下组凋落叶TN和TP的含量都会逐渐升高, 这主要是由于凋落叶质量损失的速度要快于养分释放的速度(李志安等, 2004), 这与本研究结果相一致.有研究表明, 环境因子对凋落叶分解过程中C、N和P元素含量有显著影响(岳楷等, 2015).本研究结果表明, 地上组和地下组土壤温度与凋落叶TN含量呈正相关关系, 与C/N呈负相关关系, 这是因为温度的升高提高了微生物活性, 促进了大分子含碳物质分解, 使TN含量相对升高(Mackey et al., 1996);而土壤电导率与地上组和地下组凋落叶N/P呈负相关关系, 这可能是因为电导率在一定程度上激发或抑制了微生物的活性而间接影响凋落叶元素含量动态(Gulis et al., 2006).
4.3 凋落物水解酶活性及其主控因子本研究发现, 4种水解酶活性随着分解的进行均表现出显著性差异(p < 0.01), 地上组与地下组变化趋势一致, 且二者之间无显著性差异(p>0.05).凋落叶元素含量对水解酶活性有重要的影响(Boxman et al., 1998; Sinsabaugh et al, 2002), 本研究中随着分解的进行凋落叶元素含量发生显著变化(p < 0.001), 凋落叶氮含量显著升高, 酸性磷酸酶的活性从分解前期到后期显著增强.此外, 一般在分解中期凋落物纤维素酶活性最高, 在分解前期, 可溶性含碳物质和大分子纤维素含量较高, CBH活性较高(Fioretto et al., 2007; Alarcón-Gutiérrez et al., 2010), 在第60 d达到最高值, 随着水解消耗, 相应的分解底物数量减少, CBH水解酶活性显著降低.而本研究中未发现凋落物元素含量及环境因子与βG和NAG水解酶活性之间有显著性相关, 这可能是由于随着凋落物分解水解酶之间的协同作用造成的(Sinsabaugh et al., 2010).βG和NAG水解酶在分解过程中协同其它水解酶完成分解过程, 而二者活性与凋落物元素含量不相关, 且受环境因子影响较小, 但随着分解过程的进行其活性会随其它水解酶活性的变化而显著变化.目前关于红树林凋落物分解水解酶之间的协同作用还需进一步研究.
凋落物分解过程中养分含量及土壤温度的变化对水解酶活性具有重要影响(季晓燕等, 2013a).本研究发现, 地上组和地下组秋茄凋落叶TN与AP活性均呈现显著正相关关系(p < 0.05).酶是影响凋落物分解重要的生物活性物质, 且随着凋落物类型和环境条件而发生变化(Fioretto et al, 2000; Keeler et al, 2009;季晓燕等, 2013b).许多研究表明, 增加土壤中外源氮的含量会提高土壤磷酸酶的活性(Carreiro et al, 2000; Saiyacork et al, 2002; Sinsabaugh et al, 2002), 这与本研究结论相同, 说明秋茄凋落叶的分解为红树林湿地土壤提供了大量氮的输入, 使得凋落叶表面AP水解酶活性显著提高, 促进了分解.磷酸酶作为参与磷元素循环的一种重要的水解酶, 能将磷酸盐和有机磷转化为易于被植物吸收利用的无机态磷, 其活性与土壤中有效磷含量相关.已有研究发现, 土壤AP活性与TP含量显著正相关(曹瑞等, 2016;高艳娜等, 2017), 这与本研究结果一致.对比土壤磷酸酶活性与有效磷含量的关系, 秋茄凋落叶分解过程中, 随着TP含量的升高, 凋落叶表面磷酸酶活性随之增强, 加速了分解过程的进行, 凋落物AP的活性对维持红树林生态系统磷元素的循环起到重要作用.通常,氮含量的增加会提高纤维素酶的活性(Boxman et al, 1998).也有研究报道氮含量的增加对凋落物层纤维素酶活性的影响未达到显著性水平(Andersson et al, 2004), 这与本研究结果一致.本研究中地下组AP水解酶活性与凋落叶TP含量和土壤温度具有显著的回归关系(p < 0.01), 酶活性的变化会影响凋落物的分解, 可以表征凋落物分解过程中碳、氮、磷等元素的状况, 如Waring(2013)在研究热带森林凋落物的分解时通过回归分析发现, 利用ln(BG:CBH)/ln(AP)可以对C/P进行表征;Sinsabaugh等(2010)的研究发现, ln(BG)/ln(AP)也可以表征淡水沉积物中的C/P状况.因此, 凋落物的分解往往是在水解酶作用下完成的(宋影等, 2014).
湿地土壤盐分含量的高低也会对凋落物水解酶活性产生影响, 较高的盐度会抑制酶的活性(Eliška et al., 2006).本研究以土壤电导率表征盐度(谭立山等, 2017), 结果表明, 地上组AP水解酶活性与土壤电导率呈显著正相关关系, 这可能是因为土壤较低范围的电导率容易激活凋落物水解酶的活性.目前关于盐分对红树林凋落物酶活性的研究较为少见, 其影响机制需进一步进行研究.其次, 土壤温度的变化也是影响凋落物酶活性的主要环境因子, 研究表明, 土壤AP活性与土壤温度呈显著正相关关系(曹瑞等, 2016), 这与本研究结论相同.研究发现, 增温显著降低了10~20 cm土壤深度的AP活性(Zhou et al., 2013);也有研究表明, 升温对土壤AP活性影响较小或不敏感(赵静等, 2011).总之, 升温对酶活性的影响很复杂, 可能受到酶的种类、凋落物自身的性质和季节变化等共同作用的影响.与其它陆生森林凋落物相比, 红树林作为一种热带和亚热带沿海分布的木本植物, 其凋落物分解过程中水解酶活性的变化特征需要进一步探讨.本文以秋茄为研究对象, 探究凋落叶分解过程中C、N、P含量及其化学计量比, 并结合了凋落物水解酶活性的动态变化, 这对揭示红树林生态系统凋落物分解水解酶活性机制有重要意义, 也可为红树林生态系统营养物质循环的研究提供参考性数据.
5 结论(Conclusions)1) 闽江河口秋茄凋落物量具有明显的季节变化特征, 具体表现为:8月凋落物总产量达到最高峰为141.07 g·m-2, 11月的凋落物量最少为11.77 g·m-2;不同组分所占比例不同, 表现为落叶(61.2%)>落果(23.7 %)>落枝(10.5%)>落花(4.6%);凋落物各组分也具有明显的月动态变化, 落花期出现在7—9月, 落果期为4—6月, 这是因为不同季节秋茄生长状况使生产力发生变化, 进而影响凋落物产量.此外, 不同季节秋茄各器官养分转移使凋落叶养分含量及其化学计量比发生变化, 表现为TN含量8月达到最高, TP含量1—3月显著高于其他月份, C/N在8月最低, N/P和C/P在9月最高.
2) 随着分解的进行, 地上组和地下组凋落叶养分含量变化趋势相同, 随时间变化差异性显著, 且二者TC含量、TN含量、C/N和C/P之间具有显著性差异, 凋落叶养分含量主要受到凋落物质量损失的影响, 土壤环境因子间接影响凋落叶养分含量.地上组与地下组水解酶活性变化趋势相同, 二者无显著性差异, 但随时间变化差异性显著, 水解酶活性受分解底物限制和环境因子的影响显著.
Ågren G I. 2008. Stoichiometry and nutrition of plant growth in natural communities[J]. Annual Review of Ecology Evolution and Systematics, 39(39): 153–170.
|
Alarcón-Gutiérrez E, Floch C, Ziarelli F, et al. 2010. Characterization of a Mediterranean litter by 13C CPMAS NMR:relationships between litter depth, enzyme activities and temperature[J]. European Journal of Soil Science, 59(3): 486–495.
|
Andersson M, Kjoller A, Struwe S. 2004. Microbial enzyme activities in leaf litter, humus and mineral soil layers of European forests[J]. Soil Biology and Biochemistry, 36(10): 1527–1537.
DOI:10.1016/j.soilbio.2004.07.018
|
Arreola-Lizárraga J A, Flores-Verdugo F J, Ortega-Rubio A. 2004. Structure and litterfall of an arid mangrove stand on the Gulf of California, Mexico[J]. Aquatic Botany, 79(2): 137–143.
DOI:10.1016/j.aquabot.2004.01.012
|
Attiwill P M, Adams M A. 2010. Nutrient cycling in forests[J]. New Phytologist, 124(4): 561–582.
|
Bosire J O, Dahdouh-Guebas F, Kairo J G, et al. 2005. Litter degradation and CN dynamics in reforested mangrove plantations at Gazi Bay, Kenya[J]. Biological Conservation, 126(2): 287–295.
DOI:10.1016/j.biocon.2005.06.007
|
Boxman A W, Blanck K, Brandrud T E, et al. 1998. Vegetation and soil biota response to experimentally-changed nitrogen inputs in coniferous forest ecosystems of the NITREX project[J]. Forest Ecology and Management, 101(3): 65–79.
|
曹瑞, 吴福忠, 杨万勤, 等. 2016. 海拔对高山峡谷区土壤微生物生物量和酶活性的影响[J]. 应用生态学报, 2016, 27(4): 1257–1264.
|
Carreiro M M, Sinsabaugh R L, Repert D A, et al. 2000. Microbial enzyme shifts explain litter decay responses to simulated nitrogen deposition[J]. Ecology, 81(9): 2359–2365.
DOI:10.1890/0012-9658(2000)081[2359:MESELD]2.0.CO;2
|
Chanda A, Akhand A, Manna S, et al. 2015. Mangrove associates versus true mangroves:a comparative analysis of leaf litter decomposition in Sundarban[J]. Wetlands Ecology and Management, 24(3): 1–23.
|
Chapman S B.1976.Production Ecology and Nutrient Budgets[M].London UK: Blackwell
|
Dick T M, Osunkoya O O. 2000. Influence of tidal restriction floodgates on decomposition of mangrove litter[J]. Aquatic Botany, 68(3): 273–280.
DOI:10.1016/S0304-3770(00)00119-4
|
Eliška R, Kateřina H. 2006. Wetland plant decomposition under different nutrient conditions:what is more important, litter quality or site quality[J]. Biogeochemistry, 80(3): 245–262.
DOI:10.1007/s10533-006-9021-y
|
Fenchel T, King G M, Blackburn T H. 1999. Bacterial biogeochemistry:the ecophysiology of mineral cycling[J]. Limnology and Oceanography, 44(1): 233–234.
DOI:10.4319/lo.1999.44.1.0233
|
Feller I C, Whigham D F, O'Neill J P, et al. 1999. Effects of nutrient enrichment on within-stand cycling in a mangrove forest[J]. Ecology, 80(7): 2193–2205.
DOI:10.1890/0012-9658(1999)080[2193:EONEOW]2.0.CO;2
|
Fernando S, Bandeira S O. 2009. Litter fall and decomposition of mangrove species Avicennia marina and Rhizophora mucronata in Maputo Bay, Mozambique[J]. Western Indian Ocean Journal of Marine Science, 8(2): 856–860.
|
Fioretto A, Papa S, Curcio E, et al. 2000. Enzyme dynamics on decomposing leaf litter of Cistus incanus and Myrtus communis in a Mediterranean ecosystem[J]. Soil Biology and Biochemistry, 32(13): 1847–1855.
DOI:10.1016/S0038-0717(00)00158-9
|
Fioretto A, Papa S, Pellegrino A, et al. 2007. Decomposition dynamics of Myrtus communis and Quercus ilex leaf litter:Mass loss, microbial activity and quality change[J]. Applied Soil Ecology, 36(1): 32–40.
|
高艳娜, 戚志伟, 仲启铖, 等. 2017. 长江口芦苇湿地土壤酶活性对长期模拟升温的响应[J]. 应用与环境生物学报, 2017, 23(3): 535–541.
|
Garcia-Pausas J, Casals P, Romanyà J. 2004. Litter decomposition and faunal activity in Mediterranean forest soils:effects of N content and the moss layer[J]. Soil Biology and Biochemistry, 36(6): 989–997.
DOI:10.1016/j.soilbio.2004.02.016
|
葛晓改, 肖文发, 曾立雄, 等. 2014. 三峡库区马尾松林土壤-凋落物层酶活性对凋落物分解的影响[J]. 生态学报, 2014, 34(9): 2228–2237.
|
Gulis V, Ferreira V, Graça M A S. 2006. Stimulation of leaf litter decomposition and associated fungi and invertebrates by moderate eutrophication:implications for stream assessment[J]. Freshwater Biol, 51(9): 1655–1669.
DOI:10.1111/fwb.2006.51.issue-9
|
Güsewell S. 2004. N:P ratios in terrestrial plants:variation and functional significance[J]. New Phytologist, 164(2): 243–266.
DOI:10.1111/j.1469-8137.2004.01192.x
|
Hoque M M, Kamal A H M, Idris M H, et al. 2015. Litterfall production in a tropical mangrove of Sarawak, Malaysia[J]. Acta Zoologica Lituanica, 25(2): 157–165.
|
Imgraben S, Dittmann S. 2008. Leaf litter dynamics and litter consumption in two temperate South Australian mangrove forests[J]. Journal of Sea Research, 59(1): 83–93.
|
季晓燕, 江洪, 洪江华, 等. 2013a. 模拟酸雨对亚热带三个树种凋落叶分解速率及分解酶活性的影响[J]. 环境科学学报, 2013a, 33(7): 2027–2035.
|
季晓燕, 江洪, 洪江华, 等. 2013b. 亚热带3种树种凋落叶厚度对其分解速率及酶活性的影响[J]. 生态学报, 2013b, 33(6): 1731–1739.
|
姜沛沛, 曹扬, 陈云明. 2016. 陕西省森林群落乔灌草叶片和凋落物C、N、P生态化学计量特征[J]. 应用生态学报, 2016, 27(2): 365–372.
|
金亮, 卢昌义. 2016. 秋茄中龄林和成熟林凋落物量及其动态特征[J]. 厦门大学学报(自然版), 2016, 55(4): 611–616.
|
Jr J W D, Coronado-Molina C, Vera-Herrera F R, et al. 1996. A 7 year record of above-ground net primary production in a southeastern Mexican mangrove forest[J]. Aquatic Botany, 55(1): 39–60.
DOI:10.1016/0304-3770(96)01063-7
|
Keeler B L, Hobbie S E, Kellogg L E. 2009. Effects of long-term nitrogen addition on microbial enzyme activity in eight forested and grassland sites:implications for litter and soil organic matter decomposition[J]. Ecosystems, 12(1): 1–15.
DOI:10.1007/s10021-008-9199-z
|
Lee S Y. 1989a. Litter production and turnover of the mangrove Kandelia candel (L.) druce in a Hong Kong tidal shrimp pond[J]. Estuarine Coastal and Shelf Science, 29(1): 75–87.
DOI:10.1016/0272-7714(89)90074-7
|
Lee S Y. 1989b. The importance of sesarminae crabs Chiromanthes spp.and inundation frequency on mangrove (Kandelia candel, (L.) Druce) leaf litter turnover in a Hong Kong tidal shrimp pond[J]. Journal of Experimental Marine Biology and Ecology, 131(1): 23–43.
DOI:10.1016/0022-0981(89)90009-9
|
Li L J, Zeng D H, Yu Z Y, et al. 2011. Impact of litter quality and soil nutrient availability on leaf decomposition rate in a semi-arid grassland of Northeast China[J]. Journal of Arid Environments, 75(9): 787–792.
DOI:10.1016/j.jaridenv.2011.04.009
|
Li T, Ye Y. 2014. Dynamics of decomposition and nutrient release of leaf litter in Kandelia obovata mangrove forests with different ages in Jiulongjiang Estuary, China[J]. Ecological Engineering, 73: 454–460.
DOI:10.1016/j.ecoleng.2014.09.102
|
李志安, 邹碧, 丁永祯, 等. 2004. 森林凋落物分解重要影响因子及其研究进展[J]. 生态学杂志, 2004, 23(6): 77–83.
DOI:10.3321/j.issn:1000-4890.2004.06.017 |
卢伟志, 林广旋, 王参谋, 等. 2014. 广东湛江次生与原生红树林群落碳储量与掉落物动态研究[J]. 海洋环境科学, 2014, 33(6): 913–919.
|
Mackey A P, Smail G. 1996. The decomposition of mangrove litter in a subtropical mangrove forest[J]. Hydrobiologia, 332(2): 93–98.
DOI:10.1007/BF00016688
|
Mfilinge P L, Atta N, Tsuchiya M. 2002. Nutrient dynamics and leaf litter decomposition in a subtropical mangrove forest at Oura Bay, Okinawa, Japan[J]. Trees-Struct Funct, 16(2/3): 172–180.
|
Mfilinge P L, Meziane T, Bachok Z, et al. 2005. Litter dynamics and particulate organic matter outwelling from a subtropical mangrove in Okinawa Island, South Japan[J]. Estuarine Coastal and Shelf Science, 63(1): 301–313.
|
Nga B T, Tinh H Q, Tam D T, et al. 2005. Young mangrove stands produce a large and high quality litter input to aquatic systems[J]. Wetlands Ecology and Management, 13(5): 569–576.
DOI:10.1007/s11273-004-6073-4
|
Numbere A O, Camilo G R. 2017. Mangrove leaf litter decomposition under mangrove forest stands with different levels of pollution in the Niger River Delta, Nigeria[J]. African Journal of Ecology, 55(2): 162–167.
DOI:10.1111/aje.2017.55.issue-2
|
邱明红, 岑选才, 陈毅青, 等. 2017. 森林生态系统凋落物的生产与分解[J]. 中国野生植物资源, 2017, 36(5): 45–52.
DOI:10.3969/j.issn.1006-9690.2017.05.011 |
曲浩, 赵学勇, 赵哈林, 等. 2010. 陆地生态系统凋落物分解研究进展[J]. 草业科学, 2010, 27(8): 44–51.
|
Saiyacork K, Sinsabaugh R, Zak D. 2002. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil[J]. Soil Biology and Biochemistry, 34(9): 1309–1315.
DOI:10.1016/S0038-0717(02)00074-3
|
Sánchez-Andrés R, Sánchez-Carrillo S, Alatorre L C, et al. 2010. Litterfall dynamics and nutrient decomposition of arid mangroves in the Gulf of California:Their role sustaining ecosystem heterotrophy[J]. Estuarine Coastal and Shelf Science, 89(3): 191–199.
DOI:10.1016/j.ecss.2010.07.005
|
Saiyacork K R, Sinsabaugh R L, Zak D R. 2002. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum, forest soil[J]. Soil Biology and Biochemistry, 34(9): 1309–1315.
DOI:10.1016/S0038-0717(02)00074-3
|
Salah Y M S, Scholes M C. 2011. Effect of temperature and litter quality on decomposition rate of Pinus patula, needle litter[J]. Procedia Environmental Sciences, 6(1): 180–193.
|
盛华夏.2009.九龙江口红树林湿地凋落物分解与重金属动态研究[D].厦门: 厦门大学
|
宋影, 辜夕容, 严海元, 等. 2014. 中亚热带马尾松林凋落物分解过程中的微生物与酶活性动态[J]. 环境科学, 2014, 35(3): 1151–1158.
|
Silva C A R, Lacerda L D, Ovalle A R, et al. 1998. The dynamics of heavy metals through litterfall and decomposition in a red mangrove forest[J]. Mangroves and Salt Marshes, 2(3): 149–157.
DOI:10.1023/A:1009923223882
|
Silva C A R, Sérgio R O, Ronaldo D P R, et al. 2007. Dynamics of phosphorus and nitrogen through litter fall and decomposition in a tropical mangrove forest[J]. Marine Environmental Research, 64(4): 524–534.
DOI:10.1016/j.marenvres.2007.04.007
|
Sinsabaugh R L, Carreiro M M, Repert D A. 2002. Allocation of extracellular enzymatic activity in relation to litter composition, N deposition, and mass loss[J]. Biogeochemistry, 60(1): 1–24.
DOI:10.1023/A:1016541114786
|
Sinsabaugh R L, Gallo M E, Lauber C, et al. 2005. Extracellular enzyme activities and soil organic matter dynamics for northern hardwood forests receiving simulated nitrogen deposition[J]. Biogeochemistry, 75(2): 201–215.
DOI:10.1007/s10533-004-7112-1
|
Sinsabaugh R L, Hill B H, Follstad Shah J J. 2010. Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment[J]. Nature, 462(7274): 795–798.
|
Tam N, Wong Y S, Lan C Y, et al. 1998. Litter production and decomposition in a subtropical mangrove swamp receiving wastewater[J]. Journal of Experimental Marine Biology and Ecology, 226(1): 1–18.
DOI:10.1016/S0022-0981(97)00233-5
|
谭立山, 杨平, 何露露, 等. 2017. 闽江口短叶茳芏+芦苇沼泽湿地大、小潮日土壤间隙水溶解性CH4与CO2浓度日动态[J]. 环境科学, 2017, 38(1): 52–59.
|
Twilley R R. 1985. The exchange of organic carbon in basin mangrove forests in a southwest Florida estuary[J]. Estuarine Coastal and Shelf Science, 20(5): 543–557.
DOI:10.1016/0272-7714(85)90106-4
|
Valiela I, Teal J M, Allen S D, et al. 1985. Decomposition in salt marsh ecosystems:the phases and major factors affecting disappearance of above-ground organic matter[J]. Journal of Experimental Marine Biology and Ecology, 89(1): 29–54.
DOI:10.1016/0022-0981(85)90080-2
|
Valk A G V D, Attiwill P M. 1984. Decomposition of leaf and root litter of Avicennia marina, at Westernport Bay, Victoria, Australia[J]. Aquatic Botany, 18(3): 205–221.
DOI:10.1016/0304-3770(84)90062-7
|
Wafar S, Untawale A G, Wafar M. 1997. Litter fall and energy flux in a mangrove ecosystem[J]. Estuarine, Coastal and Shelf Science, 44(1): 111–124.
DOI:10.1006/ecss.1996.0152
|
Wang C Y, Lü Y N, Wang L, et al. 2013. Insights into seasonal variation of litter decomposition and related soil degradative enzyme activities in subtropical forest in China[J]. Journal of Forestry Research, 24(4): 683–689.
DOI:10.1007/s11676-013-0405-2
|
Waring B G. 2013. Exploring relationships between enzyme activities and leaf litter decomposition in a wet tropical forest[J]. Soil Biology and Biochemistry, 64(9): 89–95.
|
Wium-Andersen S. 1981. Seasonal growth of mangrove trees in Southern Thailand.Ⅲ.Phenology of Rhizophora mucronata, Lamk.and Scyphiphora hydrophyllacea, Gaertn[J]. Aquatic Botany, 10(81): 371–376.
|
Ye Y, Chen Y P, Chen G C. 2013. Litter production and litter elemental composition in two rehabilitated Kandelia obovata, mangrove forests in Jiulongjiang Estuary, China[J]. Marine Environmental Research, 83(2): 63–72.
|
Yang J, Zhang X, Ma L, et al. 2014. Ecological stoichiometric relationships between components of Robinia pseudoacacia forest in loess plateau[J]. Acta Pedologica Sinica, 51(1): 133–142.
|
杨秀.2015.珠海市淇澳岛三种人工红树凋落物动态变化规律及凋落物分解研究[D].广州: 华南师范大学
http://d.wanfangdata.com.cn/Thesis/Y2832535 |
岳楷, 杨万勤, 彭艳, 等. 2015. 岷江上游高山森林冬季河流中凋落叶碳、氮和磷元素动态特征[J]. 应用与环境生物学报, 2015, 21(2): 301–307.
|
张川, 杨万勤, 岳楷, 等. 2016. 高山森林溪流凋落叶冬季水溶性碳含量动态[J]. 环境科学学报, 2016, 36(3): 792–801.
|
Zhang D L, Mao Z J, Zhang L, et al. 2006. Advances of enzyme activities in the process of litter decomposition[J]. Scientia Silvae Sinicae, 42(1): 105–109.
|
张乔民, 陈永福. 2003. 海南三亚河红树凋落物产量与季节变化研究[J]. 生态学报, 2003, 23(10): 1977–1983.
DOI:10.3321/j.issn:1000-0933.2003.10.004 |
赵静, 韩甜甜, 谢兴斌, 等. 2011. 酸化梨园土壤酶活性与土壤理化性质之间的关系[J]. 水土保持学报, 2011, 25(4): 115–120.
|
郑逢中, 林鹏, 卢昌义, 等. 1998. 福建九龙江口秋茄红树林凋落物年际动态及其能流量的研究[J]. 生态学报, 1998, 18(2): 113–118.
DOI:10.3321/j.issn:1000-0933.1998.02.001 |
Zheng S, Shangguan Z. 2007. Spatial patterns of leaf nutrient traits of the plants in the Loess Plateau of China[J]. Trees, 21(3): 357–370.
DOI:10.1007/s00468-007-0129-z
|
Zhou X Q, Chen C R, Wang Y F, et al. 2013. Warming and increased precipitation have differential effects on soil extracellular enzyme activities in a temperate grassland[J]. Science of the Total Environment, 444(2): 552–558.
|
Zhou H C, Tam F Y, Lin Y M, et al. 2012. Changes of condensed tannins during decomposition of leaves of Kandelia obovata in a subtropical mangrove swamp in China[J]. Soil Biology and Biochemistry, 44(1): 113–121.
DOI:10.1016/j.soilbio.2011.09.015
|
朱可峰, 廖宝文, 章家恩. 2011. 广州南沙人工红树林凋落物组成与季节变化的研究[J]. 华南农业大学学报, 2011, 32(4): 119–121.
DOI:10.3969/j.issn.1001-411X.2011.04.027 |