 2019, Vol. 39
2019, Vol. 39



2. 环境保护部环境规划院长江经济带生态环境联合研究中心, 北京 100012;
3. 昆明市环境科学研究院, 昆明 650032
2. United Center for Eco-Environment in Yangtze River Economic Belt, Chinese Academy for Environmental Planning, Beijing 100012;
3. Kunming Institute of Environmental Science, Kunming 650032
氮是陆生和水生生态系统生产力的关键营养元素, 活性氮(Reactive nitrogen, Nr)输入负荷(以下简称氮输入)升高有利于提高生态系统初级生产力, 但过量氮输入将导致区域水环境质量下降、生态系统退化和生态系统服务降低等问题(Clark et al., 2017; Seitzinger et al., 2017; Payne et al., 2017).区域氮输入的主要途径可以分为人类活动和天然固氮.在工业革命以前, 天然固氮输入是主导区域氮循环的主要来源.随着化肥施用、化石燃料燃烧和固氮农作物的种植等人类活动日益频繁, 人类活动输入已经成为全球和多数区域氮输入的主要来源(Galloway et al., 2004; Filoso et al., 2006).研究表明, 氮输入的时间变化和空间分布对区域水环境氮通量具有显著的解释能力(Chen et al., 2014;Hong et al., 2017), 是影响氮素环境过程的重要因素, 解析氮输入负荷特征对于区域氮污染控制具有重要意义.目前区域氮输入负荷研究多关注人为源评估, 对天然来源研究较少, 主要原因有两个:一是天然来源较为复杂, 目前缺乏科学的定量估算方法, 计算结果的不确定性较高(Cleveland et al., 1999);二是在人类活动较为强烈的地区, 天然来源一般占比较小, 且有下降趋势(Galloway et al., 2004).然而, 由于氮输入具有强烈的空间异质性, 在空间尺度较大的地区, 天然源输入如森林固氮也可能成为重要来源(Breemen et al., 2002; Galloway et al., 2004).因此, 在空间尺度较大的区域, 氮输入研究有必要同时考虑天然和人为来源, 从而充分反映区域氮负荷情势.相对于常规的污染源普查与调查方法, 从输入途径评估氮负荷对数据需求相对较少, 已经成为区域氮营养盐负荷研究的一种重要方法.长江经济带是我国经济社会发展和环境保护的热点区域, 覆盖11个省市, 占全国21%的国土面积, 承载了43%的人口数量和经济产出.由于经济社会发展的压力和对生态保护的日益重视, 长江经济带面临着严峻的水环境保护压力.作为长江经济带的主要水体, 长江干流的溶解态氮浓度和通量自20世纪70年代以来处于缓慢上升趋势(Li et al., 2014; Wang et al., 2014;李茂田等, 2014), 长江流域的氮剩余负荷也较世界上其他流域偏高(王佳宁等, 2016), 氮已经成为长江水环境质量的重要约束因子.由于区域氮素输入来源众多, 环境迁移转化过程和影响因素十分复杂, 受氮输入负荷、气象因子和土地利用等影响(Huang et al., 2014), 控氮成为国内外营养盐管理的难点和重点.目前, 针对长江流域的氮负荷和平衡研究相对较多(Li et al., 2014; Wang et al., 2014; Yan et al., 2010; Chen et al., 2016;王佳宁等, 2016), 但多从氮输入与环境效应响应关系角度进行研究, 且主要关注人为输入源, 针对长江经济带氮的负荷评估与解析的研究相对较少, 特别是针对长江经济带的多时空尺度分析更为缺乏.随着水环境管理科学化的需求, 解析长江经济带氮负荷特征对区域水环境管理具有十分重要的意义.
基于此, 长江经济带的氮素环境管理亟需回答以下问题:①长江经济带的区域氮输入负荷是多少?②氮输入负荷的热点地区在空间分布上具有什么特征?③氮输入负荷随时间是怎样变化的?④氮输入负荷与区域经济社会发展的关系是什么?能否科学回答以上问题直接影响到长江经济带氮素管理政策制定的可行性和科学性.因此, 本研究通过收集整理长江经济带1952—2016年连续多年的氮活动数据, 耦合人类活动氮输入和天然输入评估模型, 构建长江经济带区域氮输入负荷模型, 从而解析长江经济带氮输入负荷的时空变化特征, 以期为区域氮污染控制提供决策支持.
2 材料与方法(Materials and methods) 2.1 研究区概况长江经济带覆盖上海、江苏、浙江、安徽、江西、湖北、湖南、重庆、四川、贵州、云南等11个省市(图 1), 总面积205万km2, 占全国总面积的21%, 横跨我国东、中、西三大区域.长江经济带的地理范围以长江流域为主, 但两者边界不完全重合.长江经济带共跨越6个水资源一级区, 其中71.2%的面积位于长江区.长江经济带是我国综合实力最强、战略支撑作用最大的区域之一.2016年长江经济带常住人口达到59140万人, 地区生产总值为33.72万亿元(当年价), 分别占全国总量的43%和43%, 人均GDP为57014元·人-1.长江经济带地区生产总值占全国生产总值的比重呈现上升趋势, 从1952年占全国比重的39.2%上升到2016年的45.0%.作为我国的重要粮食和畜禽生产基地, 2016年长江经济带的粮食产量占全国总产量的37.5%, 大牲畜年底头数4091万头, 占全国的34.4%, 肉猪出栏头数34421万头, 占全国的50.2%.大规模的农业生产可能导致长江经济带非点源污染负荷较高.此外, 区域内发育的成渝、长江中游和长江三角洲3个城市群使得该地区也成为生活污染排放的热点地区.
 |
| 图 1 长江经济带地理位置 Fig. 1 Location of Yangtze River Economic Belt |
根据2006—2016年(不包括2014年)长江经济带11个省市范围内的主要国控断面水质监测结果, 长江经济带总氮的平均值为2.62 mg · L-1(标准差为2.77 mg · L-1).从空间分布看, 长江经济带各地区的氮浓度差异较大.东部的安徽省、江苏省和西部地区总氮超标严重.氮已经成为长江经济带水环境质量的重要约束因子.
2.2 区域氮输入负荷模型从活性氮来源上, 区域氮负荷可以分为人为源和天然源(Galloway et al., 2004), 前者是指由于人类生产、生活活动直接或间接带来的氮输入, 如农业生产中施用化肥;后者是指人类活动以外的因素导致的氮输入, 如闪电和天然植被的固氮作用.区域氮输入负荷用公式可以表述为:

|
(1) |
式中, TNI为区域氮输入总负荷(kg · km-2 · a-1);NANI和NNI分别为人为输入和天然输入负荷(kg · km-2 · a-1).
2.2.1 人为氮输入负荷评估针对前者本研究采用人类活动净氮输入模型来评估人类活动导致的氮输入量;人类活动净氮输入模型(Net Anthropogenic Nitrogen Input, NANI)是由美国康奈尔大学的Howarth提出(Howarth et al., 1996), 原理上属于准物料平衡法.由于该方法原理清晰、计算简便、参数获取便捷, 已经成为评估区域人为氮输入的重要方法, 在国内外营养盐评估研究中获得了广泛应用(张汪寿等, 2014; 高伟等, 2016; Hong et al., 2017).根据NANI的定义, 区域人为氮输入由食物输入、化肥施用、农作物固氮和大气NOy沉降4个组分构成, 具体计算公式如下:
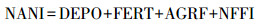
|
(2) |
式中, NANI指人类活动净氮输入量(kg · km-2 · a-1);DEPO为大气NOy沉降量(折氮)(kg · km-2 · a-1);FERT和AGRF分别表示氮肥施用量和固氮农作物的固氮量(kg · km-2 · a-1);NFFI表示通过食品或饲料贸易导致的净氮进口量(kg · km-2 · a-1).以上各组分的详细计算公式及其参数参见文献(高伟等, 2016).
2.2.2 天然氮输入负荷评估区域天然的氮输入途径主要是通过生物固氮和大气(闪电)固氮(Boyer et al., 2006), 其中闪电固氮能力较小, 仅占全球总氮输入的2%(Galloway et al., 2004), 且缺乏成熟的估算方法, 因此, 本研究仅考虑自然植被的生物固氮量.目前在大尺度上对天然生物固氮的计算方法有两种:基于天然植被面积和基于生态系统净初级生产力.其中, 基于植被面积的方法需要使用的数据为研究区多年的土地利用数据及森林和草地的固氮能力参数, 该方法数据需求小, 计算简单, 但准确度不高.本研究根据陆域生态系统的净初级生产力估算天然源输入负荷.
基于净初级生产力(NPP)的生物固氮量计算公式如下(Thornton et al., 2007;Koven et al., 2013):
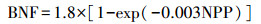
|
(3) |
式中, BNF为固氮量(g · km-2 · a-1)(以N计);NPP为净初级生产力(g · km-2 · a-1)(以C计).由于该计算方法得到的固氮量为包括农作物固氮在内的所有固氮量, 故需要扣除农作物的贡献.
NPP计算可采用气候生产力模型、生理生态过程模型和光能利用率模型.本研究根据数据可得性, 分别选用光能利用率模型GLO-PEM的计算成果和气候生产力模型Thornthiwaite.
GLO-PEM模型是基于GPP和APAR之间以光能利用率成线性关系的理论提出的, 具体方程为:
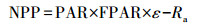
|
(4) |
式中, PAR为光合有效辐射;FPAR为植被吸收光合有效辐射比例;ε为现实光能利用率;Ra是植被自养呼吸.
Thornthiewaite模型方程(李莉等, 2014)如下:

|
(5) |

|
(6) |
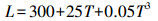
|
(7) |
式中, NPP为植被净初级生产力(g · km-2 · a-1);E为年实际蒸散量(mm);L为年平均蒸散量(mm);T为年平均气温(℃);R为年降雨量(mm).
2.3 数据来源根据氮输入负荷评估模型, 本研究需要的数据分为3类:第1类是经济社会统计数据, 第2类是下垫面植被生产力计算数据, 第3类数据是大气氮沉降数据, 各类数据的来源如下.
2.3.1 经济社会统计数据本研究使用的经济社会统计数据来源于历年中国统计年鉴、各省市统计年鉴、《中国农业统计年鉴》、各省市农业统计年鉴和《新中国六十年统计资料汇编》等, 统计时间尺度为1952—2016年.对于部分缺失数据, 采用年份回归法插补.
2.3.2 气象与初级生产力数据本研究收集了中国2000—2010年逐年NPP数据, 是基于光能利用率模型GLM-PEM计算获取的, 空间分辨率为1 km, 数据来源于中国科学院资源环境科学数据中心(http://www.resdc.cn).其他时间序列NPP根据降水和气温数据采用Thornthiewaite模型估算, 所需降水量和气温数据来自于910个国家气象站点自建站至2016年12月的八要素的日值数据, 经过年统计处理获得年降水量和年均气温.
2.3.3 氮沉降数据本研究的NOy数据来源于文献(Lamarque et al., 2010), 该数据集使用了1990s和2000s多年的数据排放源, 模拟了0.5°分辨率的全球NOy沉降量, 1952—2016年的年际变化速率参数选自文献(Lu et al., 2014), 根据基准期的NOy沉降速率和年变化速度, 插补研究期间多年的NOy沉降速率.
3 结果与讨论(Results and discussion) 3.1 人为源氮输入负荷演变特征根据人类活动净氮输入量核算结果, 长江经济带的NANI以1996年为界从1952—2016年呈现两个阶段变化特征.其中, 1952—1996年呈现显著的指数增长趋势(p < 0.01), 1997—2016年保持相对稳定的特点.1952年长江经济带的NANI为1480 Gg · a-1, 折合单位面积为721 kg · km-2 · a-1, 到1996年增长到较大值, 达到13096 Gg · a-1, 折合6380 kg · km-2 · a-1, 1952—1996年的年均增长率为5%, 随后保持相对稳定.1997—2010年虽呈增长趋势, 但基本保持在6600 kg · km-2 · a-1左右.与中国平均NANI输入相比(Han et al., 2014), 长江经济带的输入强度更高;与长江流域相比(Chen et al., 2016), 两者数量和趋势较为接近.
 |
| 图 2 1952—2016年长江经济带NANI变化 Fig. 2 Temporal change of NANI for Yangtze River Economic Belt from 1952 to 2016 |
从长江经济带的人为氮输入结构看, 化肥施用是人为活动氮输入量的主要成分, 2016年占总量的86.8%, 其次是大气氮氧化物沉降, 占总量的10.7%, 作物固氮占3.3%.化肥在氮输入中的主导地位与我国和长江流域的研究结果一致(Han et al., 2014; Chen et al., 2016).从历史变化看, 化肥施用占比呈现S型增长趋势, 增长速度最快的是1960s、1970s和1980s, 进入1980s末, 化肥施用占比呈现相对平稳的变化趋势;相对于化肥施用量, 其他3种组分的占比均呈现下降趋势, 但下降趋势进入1990s呈现递减特征, 下降幅度最高的是大气沉降和食品与饲料输入.
 |
| 图 3 1952—2016年长江经济带NANI组分变化 (DEPO、FERT、AGRF和NFFI分别为大气NOy沉降量、氮肥施用量、固氮农作物的固氮量、通过食品或饲料贸易导致的净氮进口量) Fig. 3 Components of NANI for Yangtze River Economic Belt from 1952 to 2016 (DEPO, FERT, AGRF and NFFI denotes atmospheric deposition of N, synthetic N fertilizer use, crop N fixation, and net food and feed N import respectively) |
从不同地区的NANI排放强度看, 长江经济带人为氮输入呈现显著的空间分异性特征, 总体表现为从东部到西部递减的特征, 东部的上海市、江苏省、浙江省等地区是人为氮输入强度较高的地区;中部的安徽省、湖北省和湖南省由于农业化肥使用强度高, 人为氮输入水平也较高;西部地区由于人口和产业密度相对较低, 单位面积的氮输入强度不高, 这种强度分布特征从1952—2016年总体保持稳定.但从人为氮输入总量看, 氮输入则表现为从西向东递减的反向趋势特征, 四川省、云南省、湖北省和湖南省等中西部地区人为氮输入总量占长江经济带的比重较高, 均达到10%以上, 主要原因是中西部地区土地面积较大, 农业生产强度也较高, 导致总量也呈现较高水平, 从1952—2016年, 各地区的占比变化不显著, 总体表现为东、中部氮输入开始出现下降趋势, 西部地区仍然处于上升阶段.从氮输入强度的年际变化看, 各地区从1982—2016年均表现出显著增长的趋势, 值得注意的是从2000年开始, 长江经济带的氮输入总体表现较为稳定, 呈现增长和下降并存的状态, 特别是2010—2016年期间, 7个地区的NANI强度下降, 下降的地区主要位于东部和中部, 上升的区域主要位于西部.从东、中、西部占比看, 以改革开放为时间节点, 1952—1970s末, 东部地区的NANI占比呈现上升趋势, 中部相对稳定, 西部地区表现为下降趋势;改革开放后, NANI的空间分布呈现相反的变化趋势, 东部地区占比下降, 西部有所上升, 说明改革开放后, 西部地区的人为氮输入增长速度高于东部地区, 成为长江经济带人为氮负荷增长的热点区域.导致人为氮输入向西部转移的主要原因是NANI的最大输入项—化肥输入在东部地区下降.
 |
| 图 4 1952—2016年长江经济带省级NANI空间分布 Fig. 4 Contribution of NANI in different regions of Yangtze River Economic Belt from 1952 to 2016 |
根据长江经济带多年降水、气温数据, 估算出1980—2016年的NPP变化, 采用天然氮输入计算公式, 得到长江经济带多年植被固氮量.结果表明, 长江经济带植被的多年平均固氮量为1771 kg · km-2 · a-1, 标准差为5 kg · km-2 · a-1, 年际波动较小, 最小值为1754 kg · km-2 · a-1, 最大值为1779 kg · km-2 · a-1.从时间变化趋势看, 根据Mann-Kendall趋势检验法, 从1980—2015年, 整体无显著差异(p>0.10).相对于人为输入, 植被固氮量占比较小且数值稳定, 因此, 长江经济带的植被氮固定对区域氮平衡变化的影响较小.从年际变化看, 长江经济带的年际植被固氮量的变异性有增加的趋势, 根据两期(1980—1997年和1998—2015年)的对比, 前期长江经济带的固氮量标准差为3.7 kg · km-2 · a-1, 后期上升为6.2 kg · km-2 · a-1, 上升幅度显著.扣除农作物固氮贡献后, 长江经济带其他植被的固氮量平均为763 kg · km-2 · a-1, 变化范围为761~766 kg · km-2 · a-1, 占所有植被固氮量的43%.从1980s以来, 长江经济带BNF存在一定的上升趋势, 这可能与近年来平均气温升高有关.
 |
| 图 5 1952—2016年长江经济带植被固氮量 Fig. 5 Vegetable nitrogen fixation for Yangtze River Economic Belt from 1952 to 2016 |
鉴于长江经济带植被固氮量的年际变化较小, 本研究使用多年平均值表征各地区的植被固氮能力.考虑数据质量和空间分辨率, 使用2000—2010年1 km NPP为基础数据, 计算1 km空间分辨率的固氮量分布.结果表明, 长江经济带的植被固氮量呈现显著的空间差异, 滇西北和四川西北地区的固氮量相对较低, 滇南、浙江省、安徽省等地区为高值区.从行政单元看, 浙江省是固氮强度最高的地区, 年均固氮能力为1585 kg · km-2 · a-1, 达到1500 kg · km-2 · a-1以上的其他地区包括安徽省、江西省、湖北省、湖南省和重庆市.上海市的固氮强度最低, 仅为941 kg · km-2 · a-1, 主要与上海市下垫面构成有关.上海市的城镇化程度高, 城市土地利用中城镇用地面积大、比重高, 而城镇用地由于植被覆盖度不高, 导致NPP较低, 相应的植被固氮能力也较低.剔除耕地的贡献后, 长江经济带的年均固氮能力为979 kg · km-2 · a-1.本研究使用了光能利用率模型GLO-PEM和气候生产力模型Thornthiwaite两种NPP的计算结果, 由于采用的数据与方法原理存在差异, 两种方法的计算结果不同.基于Thornthiwaite的计算结果较偏光能利用率模型GLO-PEM的计算结果大.考虑GLO-PEM计算采用的参数更为详实, 结果具有更高的空间分辨率, 本研究采用GLO-PEM对Thornthiwaite的计算结果进行了比例修正.由于缺乏研究区植被固氮研究结果, 因而难以直接与现有成果比较.考虑到本研究是基于NPP计算固氮能力, 因此, 可以通过间接比较NPP判断结果的可靠性.根据2000—2015年的NPP计算结果(李登科等, 2018), 长江经济带的NPP为608 g · km-2 · a-1(根据表格数据重新计算), 与本文结果(722 g · km-2 · a-1)较为一致.如前文所述, 由于天然固氮来源还包括闪电固氮, 本研究未考虑该项输入, 可能导致结果偏小, 但由于闪电固氮的能力较弱, 对天然固氮的影响有限.除陆地植被固氮外, 水体中(河流、湖泊)存在固氮藻类, 也具有一定的固氮能力, 但水体固氮水平相对较低, 目前湖泊蓝藻固氮的最高水平为8.9×10-9 mol · L-1 · h-1 (叶琳琳等, 2014).考虑到研究区水面覆盖比例不高, 在本研究中未加考虑.此外, 由于本研究仅剔除了耕地的NPP贡献, 但其他人工植被, 如人工林和人工草地未加以扣除, 可能导致天然氮输入结果偏大, 然而, 由于总氮输入包括天然和人为氮输入, 因此对总氮输入没有影响.
 |
| 图 6 2000—2010年基于1 km网格的长江经济带非农作物平均固氮量空间分布 Fig. 6 Non-crop nitrogen fixation rate for Yangtze River Economic Belt based on 1 km grid from averages of 2000—2010 |
根据长江经济带的人为和天然氮输入计算结果, 得到地区总氮输入量.由于天然植被固氮占比较小, 且年际波动范围较小, 所以长江经济带总氮输入量变化主要受到人为输入影响.1952—2016年长江经济带总氮输入呈现典型的S型增长趋势.1952—1970s为增长起步时期, 1980s—1990s呈现指数增长, 2000s后进入相对平稳阶段, 从2012年以后有一定的下降趋势, 主要是化肥施用和食物输入下降导致.总氮输入变化与人为输入基本一致.
从长江经济带各地区的总氮输入演变趋势看, 东部地区呈现较为典型的倒U型变化, 于1990s到达最高点, 目前处于平稳或下降阶段.相对于东部地区, 中部和西部的大部分地区仍然处于增长阶段, 特别是西部地区, 中部地区可能进入增长的顶点.从各地区的总氮输入结构看, 人为氮输入占比在研究期间呈现增长趋势, 从1980s开始在所有地区均占据主导地位, 到2016年占比为81%~99%.从各地区差异看, 东部比例相对较高, 中部次之, 西部最低.由于人口密度是NANI的重要驱动力(Han et al., 2014), 长江经济带的NANI输入强度与人口分布较为一致, 而天然输入占比相对较小, 因此, NANI空间格局成为决定NANI比例区域差异的主要因素.随着经济社会的进一步发展和土地利用变化, 人为氮输入占主导的格局不会发生改变, 将成为长江经济带氮输入的主要来源.
 |
| 图 7 1952—2016年长江经济带总氮输入量演变 (NANI、NNI和TNI分别表示人为氮输入、天然氮输入与总氮输入) Fig. 7 Temporal change of total nitrogen inputs for Yangtze River Economic Belt from 1952 to 2016 (NANI, NNI and TNI refers to anthropogenic N input, natural N input, and total N input respectively) |
 |
| 图 8 1952—2016年长江经济带各地区总氮输入量及成分演变 Fig. 8 Temporal change of total nitrogen inputs and its contribution for different regions of Yangtze River Economic Belt from 1952 to 2016 |
从长江经济带的整体来看, NANI与人均GDP呈现非线性相关, 表现出分段函数特征(图 9a), 两者关系具有一个明显的拐点, 拐点位置出现在人均GDP 9000元·人-1, 小于该值, 两者为典型的对数函数关系(R2=0.97), 高于该值, 函数关系减弱.从EKC(环境库兹涅茨曲线)理论看, 长江经济带人为氮输入可能在人均GDP为35777~36299元·人-1(拐点的95%置信区间)时出现了拐点.因此, 长江经济带整体可能已经跨越了氮输入负荷的EKC拐点, 进入经济增长与环境改善的协调发展阶段.由于地区人均GDP是持续上升的, 因此, 导致长江经济带氮输入EKC拐点出现的主要原因是NANI的下降, 根据前文分析结果, NANI降低的原因是化肥施用量的降低, 特别是东部地区下降最为显著.然而, 不同区域的EKC曲线特征有所差异.东部地区经历了较为完整的3个阶段变化, 中部地区尚处于东部地区的中间演变阶段, 经历了增长或不相关两个阶段, 而西部地区则多处于变化的初级阶段.从环境EKC理论看, 东部地区已经跨越EKC拐点, 但中部和西部地区尚未跨越.
 |
| 图 9 长江经济带NANI与人均GDP响应关系 (a.长江经济带整体, b.各地区) Fig. 9 Relationship between NANI and GDP per capita of Yangtze River Economic Belt(a.whole Yangtze River Economic Belt, b.different regions) |
1) 1952—2016年长江经济带总氮输入呈现上升、稳定和下降的阶段变化特征, 主要受到人为输入控制.各地区的总氮输入演变表现出梯级递进模式, 东部地区呈现较为典型的倒U型变化, 于1990s到达最高点, 目前处于平稳或下降阶段, 中部相对平稳, 西部的大部分地区仍然处于增长阶段.
2) 人为氮输入(NANI)以1996年为分界线呈现增长与稳定两个变化阶段特征, 其中, 1952—1996年为指数增长趋势(p < 0.01), 年均增长速度为5%, 随后保持相对稳定.从各地区差异看, 东部比例相对较高, 中部次之, 西部最低.人为输入量及其占比在空间上表现为从西到东部逐步递增的变化规律, 从1980s开始成为主导长江经济带各地区氮输入的主要来源.
3) 从人为源氮输入结构看, 化肥施用是最大来源, 2016年占长江经济带输入总量的86.8%, 化肥施用人为源比例在研究期间呈现S型增长趋势, 目前占比已进入稳定时期.
4) 长江经济带植被的多年平均固氮量为1771 kg · km-2 · a-1, 年际波动较小, 但年际变异性有增加的趋势.扣除农作物固氮贡献后, 长江经济带其他植被的固氮量平均为763 kg · km-2 · a-1, 占所有植被固氮量的43%.天然输入在长江经济带总氮负荷中数量稳定, 占比呈现下降趋势.
Boyer E W, Howarth R W, Galloway J N, et al. 2006. Riverine nitrogen export from the continents to the coasts[J]. Global Biogeochemical Cycles, 20(1): 1–91.
|
Breemen V, Boyer E W. 2002. Where did all the nitrogen go? Fate of nitrogen inputs to large watersheds in the northeastern U.S.A.[J]. Biogeochemistry, 57/58(1): 267–293.
|
Chen D, Huang H, Hu M, et al. 2014. Influence of lag effect, soil release, and climate change on watershed anthropogenic nitrogen inputs and riverine export dynamics[J]. Environmental Science & Technology, 48(10): 5683–5690.
|
Chen F, Hou L, Liu M, et al. 2016. Net anthropogenic nitrogen inputs (NANI) into the Yangtze River basin and the relationship with riverine nitrogen export[J]. Journal of Geophysical Research-Biogeosciences, 121(2): 451–465.
DOI:10.1002/2015JG003186
|
Clark C M, Bell M D, Boyd J W, et al. 2017. Nitrogen-induced terrestrial eutrophication:cascading effects and impacts on ecosystem services[J]. Ecosphere, 8(7): E01877.
DOI:10.1002/ecs2.1877
|
Cleveland C C, Townsend A R, Schimel D S, et al. 1999. Global patterns of terrestrial biological nitrogen (N2) fixation in natural ecosystems[J]. Global Biogeochemical Cycles, 13(2): 623–646.
DOI:10.1029/1999GB900014
|
Filoso S, Martinelli L A, Howarth R W, et al. 2006. Human activities changing the nitrogen cycle in Brazil[J]. Biogeochemistry, 79(1/2): 61–89.
|
Galloway J N, Dentener F J, Capone D G, et al. 2004. Nitrogen cycles:Past, present, and future[J]. Biogeochemistry, 70(2): 153–226.
DOI:10.1007/s10533-004-0370-0
|
高伟, 高波, 严长安, 等. 2016. 鄱阳湖流域人为氮磷输入演变及湖泊水环境响应[J]. 环境科学学报, 2016, 36(9): 3137–3145.
|
Han Y, Fan Y, Yang P, et al. 2014. Net anthropogenic nitrogen inputs (NANI) index application in Mainland China[J]. Geoderma, 213: 87–94.
DOI:10.1016/j.geoderma.2013.07.019
|
Hong B, Swaney D P, Mccrackin M, et al. 2017. Advances in NANI and NAPI accounting for the Baltic drainage basin:spatial and temporal trends and relationships to watershed TN and TP fluxes[J]. Biogeochemistry, 133(3): 245–261.
DOI:10.1007/s10533-017-0330-0
|
Howarth R W, Billen G, Swaney D, et al. 1996. Regional nitrogen budgets and riverine N & P fluxes for the drainages to the North Atlantic Ocean:Natural and human influences[J]. Biogeochemistry, 35(1): 75–139.
DOI:10.1007/BF02179825
|
Huang H, Chen D, Zhang B, et al. 2014. Modeling and forecasting riverine dissolved inorganic nitrogen export using anthropogenic nitrogen inputs, hydroclimate, and land-use change[J]. Journal of Hydrology, 517(5): 95–104.
|
Koven C D, Riley W J, Subin Z M, et al. 2013. The effect of vertically resolved soil biogeochemistry and alternate soil C and N models on C dynamics of CLM4[J]. Biogeosciences, 10(11): 7109–7131.
DOI:10.5194/bg-10-7109-2013
|
Lamarque J F, Bond T C, Eyring V, et al. 2010. Historical (1850-2000) gridded anthropogenic and biomass burning emissions of reactive gases and aerosols:methodology and application[J]. Atmospheric Chemistry and Physics, 10(15): 7017–7039.
DOI:10.5194/acp-10-7017-2010
|
Li H, Tang H, Shi X, et al. 2014. Increased nutrient loads from the Changjiang (Yangtze) River have led to increased harmful algal blooms[J]. Harmful Algae, 39: 92–101.
DOI:10.1016/j.hal.2014.07.002
|
李登科, 王钊. 2018. 基于MOD17A3的中国陆地植被NPP变化特征分析[J]. 生态环境学报, 2018, 27(3): 397–405.
|
李莉, 周宏飞, 包安明. 2014. 中亚地区气候生产潜力时空变化特征[J]. 自然资源学报, 2014, 29(2): 285–294.
|
李茂田, 孙千里, 王红, 等. 2014. 长江流域水库过滤器效应对入海溶解硅通量的影响[J]. 湖泊科学, 2014, 26(4): 505–514.
|
Lu C, Tian H. 2014. Half-century nitrogen deposition increase across China:A gridded time-series data set for regional environmental assessments[J]. Atmospheric Environment, 97(SI): 68–74.
|
Payne R J, Dise N B, Field C D, et al. 2017. Nitrogen deposition and plant biodiversity:past, present, and future[J]. Frontiers in Ecology and the Environment, 15(8): 431–436.
DOI:10.1002/fee.1528
|
Seitzinger S P, Phillips L. 2017. Nitrogen stewardship in the Anthropocene[J]. Science, 357(6349): 350–351.
DOI:10.1126/science.aao0812
|
Thornton P E, Lamarque J, Rosenbloom N A, et al. 2007. Influence of carbon-nitrogen cycle coupling on land model response to CO2 fertilization and climate variability[J]. Global Biogeochemical Cycles, 21: GB4018.
DOI:10.1029/2006GB002868
|
Wang J, Li X, Yan W, et al. 2014. Watershed nitrogen export model related to changing nitrogen balance and hydrology in the Changjiang River basin[J]. Nutrient Cycling in Agroecosystems, 98(1): 87–95.
DOI:10.1007/s10705-014-9598-9
|
王佳宁, 李新艳, 晏维金, 等. 2016. 基于MEA情景的长江流域氮平衡及溶解态无机氮通量:流域-河口/海湾氮综合管理[J]. 环境科学学报, 2016, 36(1): 38–46.
|
Yan W, Mayorga E, Li X, et al. 2010. Increasing anthropogenic nitrogen inputs and riverine DIN exports from the Changjiang River basin under changing human pressures[J]. Global Biogeochemical Cycles, 24(4): GB06A06.
DOI:10.1029/2009GB003575
|
叶琳琳, 张民, 孔繁翔, 等. 2014. 水生生态系统蓝藻固氮作用研究进展与展望[J]. 湖泊科学, 2014, 26(1): 9–18.
|
张汪寿, 李叙勇, 杜新忠, 等. 2014. 流域人类活动净氮输入量的估算、不确定性及影响因素[J]. 生态学报, 2014, 34(24): 7454–7464.
|