 2020, Vol. 40
2020, Vol. 40



2. 福建师范大学地理研究所, 福州 350007;
3. 福建师范大学福建省亚热带资源与环境重点实验室, 福州 350007
2. Key Laboratory of Humid Subtropical Eco-geographical Process(Fujian Normal University), Ministry of Education, Fuzhou 350007;
3. Fujian Provincial Key Laboratory for Subtropical Resources and Environment, Fujian Normal University, Fuzhou 350007
湿地土壤是供给植物生长发育所需营养物质的重要储库.湿地植物除需从土壤中吸收大量营养元素来满足自身生长发育外, 亦需要吸收一定量的微量元素以维持某些特定生理生态过程的进行(祝贺等, 2016).钒(V)是植物生长发育过程中不可或缺的微量元素, 其在生物固氮以及植物体内多种酶促反应过程中均发挥着十分重要的作用.适量的V不但可促进湿地植物的固氮作用, 而且亦能够促进植物对铁的吸收与利用, 促进叶绿素的合成, 产生希尔反应(王云等, 1995;Nawaz et al., 2018);而过量的V又会抑制湿地植物根系细胞膜上的多种ATP酶, 导致植物发育不良、矮化及生物量降低, 同时也会减少植物对钙和磷酸盐等营养元素的吸收(矫旭东等, 2008).湿地植物对V的生物富集程度不但可反映其在土壤中的供给状况, 而且亦可在一定程度上指示其在湿地系统中的生物地球化学行为.河口湿地作为滨海湿地的一个重要湿地类型, 处于陆地和海洋交汇处, 河流的径流物质在河口区输送入海时, 受潮流与径流咸淡水相互作用的影响, 引起介质的物理及化学条件(如粒度、pH、Eh、盐度和水位等)等急剧变化, 加之水动力作用强烈, 泥沙冲淤变化频繁以及高强度人类活动的影响, 导致河口区元素的富集规律及生物地球化学过程非常复杂(张晓晓, 2010), 对其进行生态修复或污染治理对于保障海岸带地区的生态安全具有重要意义.目前, 国内外关于河口湿地植物-土壤系统的元素研究主要集中在对碳、氮、磷和硫(Korol et al, 2016; Herbert et al, 2018; Wan et al, 2020; He et al, 2020)等大量或中量营养元素以及铁、锰、铅、铜、锌、铬、镍等重金属(陈权和马克明, 2017;Amr et al, 2020)探讨上, 而关于V等微量营养元素的相关研究还较为薄弱.
闽江河口位于中亚热带和南亚热带的过渡区, 是我国最典型的亚热带河口之一.闽江河口湿地是闽江与东海相互作用形成的特殊生态类型, 受河口水动力、海洋潮汐及植被演替等多因素影响, 河口湿地的环境条件极为复杂(何涛等, 2018), 由此可能直接或间接影响到湿地植物-土壤系统中V的赋存形态、分布特征及其生物地球化学行为.湿地植物的空间扩展实质上反映了不同物种之间对光、水分和养分等环境资源以及生存空间的竞争, 而竞争是塑造植物形态特征和群落结构的重要动力(秦先燕等, 2010;王华等, 2019).短叶茳芏(Cyperus malaccensis)是闽江河口分布最为广泛的原生植被之一, 而芦苇(Phragmites australis)是本地入侵物种, 它是大约30~40年前由闽江中上游地区逐渐扩散至闽江河口鳝鱼滩湿地.现有研究表明, 闽江河口芦苇与短叶茳芏的竞争能力相当, 二者的空间扩展是双向过程, 但芦苇具有相对竞争优势(Li et al., 2020), 由此导致短叶茳芏湿地在空间尺度上逐渐被芦苇湿地扩展, 表现为短叶茳芏湿地面积缓慢萎缩, 而芦苇与短叶茳芏的交错带变宽(宽度达100~150 m), 且其面积呈逐年扩大趋势.Ewanchuk等(2004)的研究指出, 湿地植被间的空间扩展不仅可对湿地沉积环境及土壤理化性质产生重要影响, 而且亦会导致植物群落结构发生重大改变, 进而影响生态系统的物质循环、能量流动及其稳定性.相对于空间扩展前而言, 芦苇与短叶茳芏空间扩展过程中的土壤理化性质以及植物对养分的吸收利用策略等均发生了较为明显的改变(何涛等, 2018).
当前, 关于闽江河口湿地植物-土壤系统元素的研究主要集中于对碳、氮、磷及硫的探讨上, 而对微量元素的研究相对较少, 且现有关于微量元素的研究主要侧重于重金属的赋存及其生态风险(蔡海洋等, 2011;Sun et al, 2019), 而关于V等微量营养元素赋存及其生物富集的相关研究还鲜有报道.特别是, 不同植被空间扩展影响下V的生物富集特征究竟有何变化?尚不明确.对此问题的探讨, 有助于揭示湿地植物间的微量营养元素竞争机制, 为此, 本研究选择闽江河口鳝鱼滩芦苇与短叶茳芏空间扩展前的芦苇湿地、短叶茳芏湿地以及二者空间扩展过程中的交错带湿地为研究对象, 探讨了湿地植被空间扩展对V生物富集特征的影响, 研究结果为该区的湿地生态保育提供重要的科学依据.
2 材料与方法(Materials and methods) 2.1 研究区概况闽江河口地处我国东南部, 气候暖热潮湿, 雨热同期, 年平均气温为19.6 ℃, 年降水量超过1350 mm.本研究区位于闽江河口鳝鱼滩(119°34′12″~119°40′40″ E, 26°00′36″~26°03′42″ N)(图 1), 潮汐为典型的半日潮, 主潮沟由西北向东南方向延伸, 水文过程较为复杂.该区湿地土壤类型为滨海盐土, 主要植被包括短叶茳芏(C. malaccensis)、芦苇(P. australis)、互花米草(Spartina alterniflora)和扁穗莎草(C. compressus), 植被群落由陆向海整体呈条带状分布.
 |
| 图 1 研究区域及研究样地示意图 Fig. 1 Sketch of the study region and sampling sites |
2015年7月, 基于时空互代法在鳝鱼滩西北部由陆向海方向布设A、B两条重复样带.在每条样带上分别设置芦苇湿地(P. australis, 记为PA)、短叶茳芏湿地(C. malaccensis, 记为CM)以及两种植物空间扩展形成的交错带湿地(P. australis-C. malaccensis, 记为EM;PA′和CM′分别表示交错带中的芦苇和短叶茳芏)3个典型研究样地(图 1).地上生物量采用收获法, 即在每个样地内随机选取3个50 cm×50 cm的样方, 用剪刀沿地面割下植物地上部分, 并同步测定植物的株高和密度.地下生物量采用挖掘法, 即将样方内的植物根系全部挖出, 并将其放入细纱网袋中冲洗干净.在上述研究样地内, 采用土壤柱状采样器进行不同样地土壤样品的采集, 采集深度为60 cm, 每10 cm一层, 每个样地采集3个柱样作为重复, 共采集108个样品.不同样地表层土壤的理化性质见表 1.
| 表 1 不同湿地表层土壤理化性质 Table 1 Physical and chemical properties of topsoils in different marshes |
将采集的植物样品在实验室内分离为根、茎、叶和立枯体, 待清洗干净后放入烘干箱中杀青烘干, 称量后磨碎装袋待测.将采集的土壤样品于实验室自然风干, 去除杂物研磨过100目筛后装袋待测.准确称取0.0400 g土壤样品于高压消解罐的Teflon容器中, 加入HF和HNO3(3:1)2 mL, 置于烘箱内150 ℃加热12 h;待样品冷却后加入0.25 mL HClO4, 于150 ℃电热板上蒸至近干, 加入2 mL高纯水和1 mL HNO3, 放入烘箱内150 ℃回溶12 h;消解好的样品用高纯水定容至10 mL待测.称取0.2000 g植物样品, 加入2 mL 65%的HNO3和1 mL30%的H2O2消解后, 残渣用去离子水稀释至10 mL待测.土壤和植物的V含量采用美国Thermo Fisher公司的X-SerieⅡ型ICP-MS质谱仪进行测定, 以Rh和Re为内标对实验过程进行质量监控, 平行测试RSD<5%, 实验用水均为18.2 MΩ高纯水.为保证实验方法的可靠性, 同步测定国家标准水系沉积物参比物质(GSD-9)、空白和平行样进行质量控制.土壤有机质含量采用土壤养分分析仪(TFW-Ⅲ)测定;土壤粒度采用Master Sizer 2000激光粒度分析仪测定, 按照国际制分类.植物和土壤样品采集的同时, 采用HACH-sensION3和ECTestr11+分别原位测定土壤pH和EC.
2.4 相关指标计算植物V含量的根/茎比(R/S)、根/叶比(R/L)和茎/叶比(S/L)采用下式计算(Dahmani-Muller, 2000):
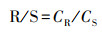
|
(1) |
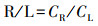
|
(2) |
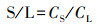
|
(3) |
式中, CR、CS和CL分别为根、茎和叶的V含量(mg·kg-1).
植物不同器官的V生物富集系数(bioconcentration factors, BCF)可用各器官V含量与土壤V含量的比值来表征(Sun et al, 2015), 即:
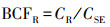
|
(4) |
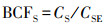
|
(5) |
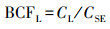
|
(6) |
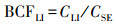
|
(7) |
式中,CR、CS、CL同上,CLI为立枯体的V含量(mg·kg-1);CSE为土壤的V含量(mg·kg-1).
2.5 数据处理与分析利用Origin8.0软件对数据进行计算和作图;采用SPSS20.0软件对数据进行单因素方差关分析、Pearson相关分析和逐步线性回归分析, 显著性水平设定为p=0.05;运用Canono5.0软件对数据进行主成分分析.
3 结果与分析(Results and analyses) 3.1 空间扩展对湿地土壤V含量分布的影响芦苇与短叶茳芏空间扩展影响下, 不同湿地土壤中的V含量在0~40 cm各土层均表现为芦苇湿地>交错带湿地>短叶茳芏湿地, 在40~50 cm土层表现为交错带湿地>芦苇湿地>短叶茳芏湿地, 而在50~60 cm土层表现为短叶茳芏湿地>交错带湿地>芦苇湿地(图 2).其中, 芦苇湿地与短叶茳芏湿地在20~30 cm土层存在显著差异(p<0.05).在垂直方向上, 芦苇湿地和交错带湿地土壤的V含量垂直变化特征较为一致, 且均于40~50 cm土层出现较高值.短叶茳芏湿地0~30 cm土层的V含量变化趋势与交错带湿地较为相近, 而在30~60 cm土层则呈相反变化特征(图 2).比较而言, 芦苇湿地与短叶茳芏湿地(p=0.044)以及交错带湿地与短叶茳芏湿地均存在显著差异(p=0.039).就平均状况来说, 芦苇湿地、交错带湿地和短叶茳芏湿地土壤的V含量均值分别为(107.59±5.10)、(107.82±9.48)和(99.20±3.90) mg·kg-1, 变异系数分别为47.44%、87.94%和39.37%.与短叶茳芏湿地相比, 交错带湿地土壤的V含量增加了8.69%;但与芦苇湿地相比, 其V含量仅增加了0.21%.
 |
| 图 2 不同湿地土壤钒(V)含量垂直分布 (a, b表示不同湿地V含量之间的差异, 字母不同表示在p<0.05水平上存在显著差异) Fig. 2 Distribution of vanadium (V) contents in the profile of different marshes |
芦苇与短叶茳芏空间扩展过程中, 纯群落芦苇根和叶的V含量均低于交错带芦苇, 而其茎和立枯体的V含量均要高于交错带芦苇(图 3), 且二者叶的V含量存在显著差异(p=0.007).纯群落短叶茳芏根和叶的V含量均高于交错带短叶茳芏, 而其茎和立枯体的V含量均低于交错地短叶茳芏, 且二者叶的V含量存在极显著差异(p=0.005).另外, 交错带芦苇根、茎、叶和立枯体的V含量均低于交错带短叶茳芏, 且二者叶的V含量存在显著差异(p=0.018).纯群落芦苇根、叶和立枯体的V含量均低于纯群落短叶茳芏, 而其茎的V含量要高于纯群落短叶茳芏, 且二者根(p=0.013)和叶(p<0.01)的V含量均存在显著差异(图 3).
 |
| 图 3 不同湿地植被各部分的V含量 (a, b表示不同湿地相同植被V含量之间的差异, m, n表示不同植被相同器官V含量之间的差异, 字母不同表示在p<0.05水平上存在显著差异) Fig. 3 Vanadium (V) contents in organs of different marsh plants |
纯群落芦苇中V的R/S、R/L和S/L均高于交错带芦苇, 而纯群落短叶茳芏除R/S较高外, 其R/L和S/L均低于交错带短叶茳芏(图 4).另外, 交错带短叶茳芏除S/L较低外, 其R/S和R/L均高于交错带芦苇.纯群落芦苇除R/S较低外, 其R/L和S/L值均明显高于纯群落短叶茳芏.整体而言, 纯群落芦苇和交错带芦苇的R/S、R/L和S/L均大于1, 而纯群落短叶茳芏和交错带短叶茳芏的R/S和R/L均大于1.
 |
| 图 4 不同器官V的R/S、R/L和S/L比值 Fig. 4 Root/stem (R/S), root/leaf (R/L) and stem/leaf (S/L) ratios of V in organs of different plants |
芦苇与短叶茳芏空间扩展影响下, 纯群落芦苇根和叶的BCF值均低于交错带芦苇, 而其茎和立枯体的BCF值均高于交错带芦苇;纯群落短叶茳芏根和叶的BCF值均高于交错带短叶茳芏, 而其茎和立枯体的BCF值均低于交错带短叶茳芏(图 5).另外, 交错带短叶茳芏根、茎、叶和立枯体的BCF值均高于交错带芦苇, 而纯群落芦苇除茎的BCF值较高外, 其根、叶和立枯体的BCF值均低于纯群落短叶茳芏(图 5).
 |
| 图 5 不同湿地植物各部分V的富集系数 Fig. 5 Bioconcentration factors (BCF) of V in organs of different marsh plants |
土壤中的微量元素主要来源于成土母质, 而大气沉降、人为活动也输入了一定量的微量元素, 成土母质和成土过程是影响土壤V含量的首要因素.已有研究表明, 在V含量高的母岩上发育的土壤, 其V含量相应也较高(汪金舫等, 1994).闽江河口的湿地土壤主要来源于闽江水系带来的泥沙以及河口的泥沙淤积, 其土壤母质主要为红壤、黄壤及滨海盐土.研究区表层土壤的V含量为101.11 mg·kg-1, 高于福建省土壤V含量的背景值(78.3 mg·kg-1)(陈振金, 1992), 但低于我国陆地表面的V含量背景值(112 mg·kg-1)(聂兰仕, 2011), 说明闽江河口湿地土壤存在V的富集过程.通常用富集系数来表征元素的富集程度, 其值为研究区的元素含量与该研究区的背景值(地壳元素丰度)之比(祝贺等, 2016).研究发现, 闽江河口湿地土壤的富集系数为1.29, 整体较低, 为弱程度的富集.另外, 本研究区域位于福建闽江河口湿地国家级自然保护区内, 保护区实行最严格的一级管控政策, 禁止一切形式的开发建设活动.因此, 本研究区不受外来污染(如工农业废水和生活污水排放)的影响.
本研究表明, 芦苇与短叶茳芏空间扩展影响下不同湿地土壤的V含量在0~40 cm土层整体表现为芦苇湿地>交错带湿地>短叶茳芏湿地, 这一方面可能与不同湿地受潮汐影响程度差异导致主要环境因子(pH、EC)发生变化有关, 另一方面可能与空间扩展影响下土壤颗粒组成、植物根系分布深度以及其对V的吸收利用发生改变有关.已有研究表明, 沉积物中水的较强流动性有利于金属离子的迁移(Sun et al, 2015), 而伴随着水在沉积物中的强烈流动, 水中的盐度还可通过与盐阴离子的络合以及阳离子与金属离子的离子交换而促进金属离子的迁移(Li et al, 2011).本研究中, 芦苇湿地距海最远, 交错带湿地次之, 而短叶茳芏湿地距海最近, 所以短叶茳芏湿地土壤中较高的EC值(表 1)极有可能增强V的迁移性, 导致其含量较低(图 2);而芦苇湿地和交错带湿地土壤中较低的EC值(表 1)可能导致V的迁移性相对较弱, 导致其含量相对较高(图 2).另有研究表明, V在表生带氧化条件下, V3+可转化为V5+并形成易溶于水, 且迁移能力强的VO43-络阴离子可导致V的富集, 而这种转化能力往往随pH值的升高而增强;但在还原环境或滞水环境中, 则不利于V的富集(曾英等, 2004).由于芦苇湿地受潮汐影响最弱, 而短叶茳芏湿地受潮汐影响最为明显, 这就导致前者处于滞水厌氧环境中的时间较短, 而后者所处滞水厌氧环境中的时间相对较长, 由此使得芦苇湿地土壤中V的富集程度可能要高于短叶茳芏湿地.正是由于上述淹水条件导致的氧化还原条件的差异, 使得短叶茳芏湿地0~40 cm土层的V含量要低于交错带湿地和芦苇湿地.相关分析表明, 短叶茳芏湿地土壤的V含量与pH呈显著负相关(p<0.05).由于短叶茳芏湿地土壤的pH要高于交错带湿地(表 1), 故其整体不利于土壤中V的富集.多元逐步线性回归分析进一步表明, 交错带湿地土壤的V含量(y1)主要受EC(x1)的影响(y1=13.571x1+61.788, R2=0.431, p=0.023), 而短叶茳芏湿地(y2)的V含量主要受黏粒(x1)和pH(x2)的影响(y2=-2.346x1-34.594x2+348.030, R2=0.89, p=0.017), 这一结果与上述分析基本吻合.另有研究表明, 芦苇湿地土壤的有机质集中分布在0~50 cm土层(吕国红等, 2006), 而0~50 cm也是本研究中芦苇根系的集中分布区.Jobbagy等(2000)研究表明, 植物根系的分布直接影响土壤有机质的垂直分布, 因为大量的死根腐解为土壤提供了丰富的有机组分.此外, 有机质可以形成络合物, 与V结合, 并将V还原到较低的价态, 从而降低V的生物利用度和毒性.一般而言, 富含有机质的土壤中V含量较高(Gu et al, 2011).本研究中, 短叶茳芏的根系大多分布于0~30 cm土层, 无论是在纯群落还是在交错带, 短叶茳芏和芦苇的V含量及BCF值均以根部最高, 且短叶茳芏根的V含量要高于芦苇(图 3、图 5), 而这可能也是导致芦苇湿地0~40 cm土层的V含量高于交错带湿地和短叶茳芏湿地的一个重要原因.本研究亦表明, 芦苇湿地和交错带湿地土壤的V含量垂直变化特征较为一致, 并均于40~50 cm土层出现较高值.由于细颗粒对V具有较强的吸附作用, 故湿地土壤的质地特别是粘粒含量可影响V的可萃取性或可浸出性.汪金舫等(1994)研究表明, 土壤全V和可溶态V含量与粘粒含量均存在显著正相关关系.本研究中, 芦苇湿地和交错带湿地土壤的黏粒含量均较高, 特别是交错带湿地的黏粒含量在40~50 cm土层取得最高值(16.44%), 由此可能导致其在该土层的V含量出现较高值.另有研究表明, 芦苇具有十分发达的根系(根系深度可达120 cm), 而发达的根系对养分具有较强的吸收作用, 且其死亡后分解归还养分的能力也较强(王为东等, 2010).本研究中, 短叶茳芏的根系大多分布于0~30 cm土层, 而芦苇根系大多分布于0~60 cm土层, 所以交错带湿地土壤受到芦苇根系的影响可能更为明显, 而这可能也是导致交错带湿地土壤的V含量垂直变化与芦苇湿地相似的一个重要原因.
本研究还表明, 芦苇与短叶茳芏的空间扩展增加了交错带湿地土壤的V含量, 其均值相比短叶茳芏湿地增加了8.69%, 而相比芦苇湿地仅增加了0.21%, 原因可能主要有两方面:一是与空间扩展过程中湿地土壤颗粒组成的变化有关.在芦苇和短叶茳芏形成的交错带, 二者因高度差异, 芦苇占据高处空间, 而短叶茳芏占据低处空间, 这种高低空间搭配(图 6a)以及二者较高的密度(图 6a)导致其对悬浮颗粒物的拦截能力相比纯群落可能更强.本研究中, 交错带湿地表层土壤中的黏粒和粉粒含量相对于芦苇湿地增加了11.8%, 而相对于短叶茳芏湿地升高了29.1%, 表明两种植物空间扩展增加了土壤中的黏粒和粉粒含量(表 1).交错带湿地土壤中细颗粒占比以及有机质含量的增加(表 1), 一方面可提高对V的吸附能力;另一方面大量的V养分亦可随细颗粒被拦截而输入到湿地土壤中, 由此导致其具有较高的V含量.二是与空间扩展过程中两种植被因存在对养分、光照等资源的竞争使得二者生态特征均发生较大变化有关.据图 6可知, 空间扩展影响下交错带芦苇的株高和生物量较纯群落均降低, 而密度有所增加;交错带短叶茳芏的生物量和密度较纯群落均降低, 而株高有所增加.可能正是由于交错带芦苇和短叶茳芏的地上、地下生物量相对于纯群落均明显降低, 使二者对其土壤中V养分的吸收能力降低, 从而导致交错带土壤中的V养分含量较高.
 |
| 图 6 芦苇与短叶茳芏空间扩展过程中植被生态特征 Fig. 6 Ecological traits of Phragmites australis and Cyperus malaccensis in the process of their spatial expansion |
为进一步明确影响芦苇与短叶茳芏空间扩展过程中湿地土壤V含量分布的关键因素, 对不同湿地土壤的V含量、环境因子以及植被生态特征进行了主成分分析(图 7).结果表明, 芦苇湿地提取的两个主成分(PCA1和PCA2)累计贡献率为99.66%.其中, PCA1的贡献率为98.99%, 可看作是植被生态特征(株高、生物量、密度)的代表;PCA2的贡献率为0.67%, 可看作是颗粒组成(粉粒、砂粒)的代表.交错带湿地提取的两个主成分(PCA1和PCA2)累计贡献率达96.87%.其中, PCA1的贡献率为82.77%, 可看作是植被生态特征(生物量)的代表;PCA2的贡献率为14.10%, 也可看作是植被生态特征(株高、密度)的代表.短叶茳芏湿地提取的两个主成分(PCA1和PCA2)累计贡献率达99.81%.其中, PCA1的贡献率为99.45%, 可看作是植被生态特征(生物量)和EC的代表;PCA2的贡献率为0.36%, 可看作是颗粒组成(黏粒和粉粒)的代表.上述结果与前述分析整体一致, 即芦苇与短叶茳芏的空间扩展主要通过改变湿地土壤颗粒组成、EC以及植物生物量、株高和密度来影响土壤的V含量分布.
 |
| 图 7 不同湿地土壤V含量与环境因子及植物生态特征主成分分析 (注:Sand, 砂粒;Silt, 粉粒;Clay, 黏粒;EC, 电导率;Moisture, 含水量;BD, 容重;Destiny, 密度;Height, 株高;AbovBiom, 地上生物量;UndrBiom, 地下生物量;SOM, 有机质;RootRati, 根冠比.) Fig. 7 Principal component analyses for V contents in soils, environmental factors and plant ecological traits in different marshes |
本研究表明, 无论是纯群落还是交错带, 芦苇和短叶茳芏的V含量及BCF值均以根部最高, 且短叶茳芏根的V含量要高于芦苇.已有研究发现, 当V存在于生长介质中时, 生物富集是大多数植物的主要适应策略, 且V主要集中在根部(Saco et al, 2013; Hou et al, 2014), 其富集量大约是地上部分的2~1000倍(Aihemaiti et al, 2020).另有研究表明, V主要通过根细胞壁极性化合物的螯合和络合而在根组织中与钙形成稳定的化合物, 这就降低了游离V离子的浓度和迁移性, 由此可能导致植物地上部分的V累积水平较低(Kaplan et al, 1990).本研究中, 不同植物的R/S和R/L均大于1(图 4), 说明上述机制可能是导致不同植物根中V富集水平较高但其向地上器官转移V较低的重要原因.
本研究还表明, 交错带芦苇根、叶的V含量和BCF值相比纯群落均升高, 而茎和立枯体的V含量和BCF值均降低.与之不同, 交错带短叶茳芏根、叶的V含量和BCF值相比纯群落均降低, 而茎和立枯体的V含量和BCF值均显著增加.导致空间扩展过程中芦苇与短叶茳芏的V富集呈现出相反变化特征的原因可能与上述二者竞争过程中植被生态特征改变以及其对不同养分(包括V)的竞争利用有关.本研究中, 相对于纯群落, 交错带芦苇的生物量、株高以及R/S、R/L、S/L均显著降低, 而密度有所增加(图 6), 说明其生存空间在竞争中受到挤压, 且V等养分的供应亦可能受到一定限制.已研究表明, 在养分受限的生境中, 植被占据竞争优势而成功侵入往往依赖于对有限资源的保守利用(Sardans et al, 2012).王华等(2018)的研究表明, 芦苇主要通过对有限养分的保守利用而在空间扩展过程中获得相对竞争优势.因此, 在生存空间及养分限制条件下, 本研究中的交错带芦苇极有可能通过显著降低地下生物量以及S/L的方式将V大量富集到根部, 并将其优先转运至光合器官(叶)的方式来保持相对竞争优势.与纯群落短叶茳芏相比, 交错带短叶茳芏的生物量、密度和R/S均显著降低, 而株高和R/L、S/L有所增加(图 6), 说明交错带短叶茳芏为缓解来自芦苇的竞争压力, 极有可能通过降低R/S、提高S/L等方式将富集到根中的V大量转移到茎中(茎为短叶茳芏地上生物量的主体, 占94.3%)以抗衡来自芦苇的竞争压力, 由此导致其茎中的V含量显著增加, 而根中的V含量显著降低.
本研究亦表明, 交错带芦苇不同器官的V含量及BCF值均低于交错带短叶茳芏, 这主要与二者在空间扩展的竞争环境中对V的吸收利用与转运的差异有关.本研究中, 交错带芦苇的地上生物量要高于交错带短叶茳芏, 而其地下生物量则低于交错带短叶茳芏(图 6b), 加之交错带短叶茳芏R/S和R/L均高于交错带芦苇(图 4), 说明交错带短叶茳芏地下器官对V的富集能力相比交错带芦苇更高.另有研究表明, 芦苇为C3为植物, 而短叶茳芏为C4植物, 其叶绿体对CO2的亲和力要强于C3植物, 而适量的V又可促进叶绿素的合成以及提高对氮、磷、钾、钙等必需营养元素的吸收与转运(Yang et al., 2018).本研究中, 交错带芦苇的R/S、R/L和S/L均大于1;与之不同, 交错带短叶茳芏的R/S和R/L均大于1, 而S/L小于1(图 4), 说明两种植物尽管转移到地上部分的V均较低, 但交错带短叶茳芏将V转运至茎、叶的比例相对较高, 从而更有利于对V的吸收利用.另外, 由于亚热带河口的高温时间长, 而当温度达到一定范围时, 芦苇便出现“光合午休”现象(何涛等, 2018).由于本研究为夏季, 所以在与短叶茳芏的竞争过程中, 芦苇“光合午休”现象的产生在一定程度上也降低了其对V的富集能力.通过上述对不同湿地植被V的富集及转移的对比研究发现, 交错带短叶茳芏可用于未来受V污染湿地修复的备选物种.
5 结论(Conclusions)1) 空间扩展影响下不同湿地土壤的V含量在0~40 cm土层均表现为芦苇湿地>交错带湿地>短叶茳芏湿地;芦苇湿地和交错带湿地土壤的V含量垂直变化特征较为一致, 并均于40~50 cm土层出现较高值.
2) 芦苇与短叶茳芏的空间扩展整体增加了交错带湿地土壤的V含量, 其均值相比短叶茳芏湿地增加了8.69%, 而相比芦苇湿地仅增加0.21%;空间扩展主要通过改变湿地土壤的颗粒组成、EC以及植物的生物量、株高和密度来影响土壤的V含量分布.
3) 空间扩展过程中, 芦苇与短叶茳芏的V生物富集特征呈相反变化, 这与二者竞争过程中植被生态特征改变以及其对V的竞争利用方式有关.
4) 不同湿地植被均以根的V富集能力最强, 且交错带短叶茳芏的V生物富集能力要高于交错带芦苇, 这与二者在竞争生境中对V的吸收利用与转运的差异有关.交错带短叶茳芏可用于未来受V污染湿地修复的备选物种.
5) 芦苇与短叶茳芏为应对空间扩展过程中的竞争可能采取不同的V富集策略, 前者通过将富集到根中的V优先转运至光合器官(叶)的方式来保持相对竞争优势, 而后者通过将富集到根中的V大量转移到地上主体(茎)的方式来抗衡来自芦苇的竞争.
Aihemaiti A, Gao Y C, Meng Y, et al. 2020. Review of plant-vanadium physiological interactions, bioaccumulation, and bioremediation of vanadium-contaminated sites[J]. Science of the Total Environment, 712: 135637. DOI:10.1016/j.scitotenv.2019.135637 |
Amr E, Keshta, Kamal H, et al. 2020. Sediment clays are trapping heavy metals in urban lakes:An indicator for severe industrial and agricultural influence on coastal wetlands at the Mediterranean coast of Egypt[J]. Marine Pollution Bulletin, 151: 110816. DOI:10.1016/j.marpolbul.2019.110816 |
蔡海洋, 曾六福, 方妙真, 等. 2011. 闽江口塔礁洲湿地重金属的分布特征[J]. 福建农林大学学报(自然科学版), 40(3): 285-289. |
陈权, 马克明. 2017. 互花米草入侵对红树林湿地沉积物重金属累积的效应与潜在机制[J]. 植物生态学报, 41(4): 409-417. |
陈振金, 陈春秀, 刘用清, 等. 1992. 福建省土壤元素背景值及其特征[J]. 中国环境监测, 8(3): 107-110. |
Dahmani-Muller H, van Oort F, Gélie B, et al. 2000. Strategies of heavy metal uptake by three plant species growing near a metal smelter[J]. Environmental Pollution, 109(2): 231-238. DOI:10.1016/S0269-7491(99)00262-6 |
Ewanchuk P J, Bertness M D. 2004. Structure and organization of a northern New England salt marsh plant community[J]. Journal of Ecology, 92(1): 72-85. DOI:10.1111/j.1365-2745.2004.00838.x |
Gu H H, Qiu H, Tian T, et al. 2011. Mitigation effects of silicon rich amendments on heavy metal accumulation in rice (Oryza sativa L.) planted on multi-metal contaminated acidic soil[J]. Chemosphere, 83(9): 1234-1240. DOI:10.1016/j.chemosphere.2011.03.014 |
He T, Sun Z G, Hu X Y, et al. 2020. Effects of Spartina alterniflora invasion on spatial and temporal variations of total sulfur and inorganic sulfur fractions in sediments of salt marsh in the Min River estuary, southeast China[J]. Ecological Indicators, 113: 106253. DOI:10.1016/j.ecolind.2020.106253 |
何涛, 孙志高, 李家兵, 等. 2018. 闽江河口芦苇与短叶茳芏空间扩展植物-土壤系统硫含量变化特征[J]. 生态学报, 38(5): 1607-1618. |
Hou M, Lu C, Wei K. 2014. Accumulation and Speciation of Vanadium in Lycium Seedling[J]. Biological Trace Element Research, 159(1/3): 373-378. |
Herbert E R, Schubauer-Berigan J, Craft C B. 2018. Differential effects of chronic and acute simulated seawater intrusion on tidal freshwater marsh carbon cycling[J]. Biogeochemistry, 138(2): 137-154. DOI:10.1007/s10533-018-0436-z |
Jobbagy E G, Jackson R B. 2000. The vertical distribution of soil organic carbon and it's relation to climate and vegetation[J]. Ecological Applications, 10(2): 423-436. DOI:10.1890/1051-0761(2000)010[0423:TVDOSO]2.0.CO;2 |
矫旭东, 滕彦国. 2008. 土壤中钒污染的修复与治理技术研究[J]. 土壤通报, 39(2): 448-452. |
Kaplan D I, Sajwan K S, Adriano D C, et al. 1990. Phytoavailability and toxicity of beryllium and vanadium[J]. Water, Air and Soil Pollution, 53: 203-212. |
Korol A R, Changwoo Ahn, Noe G B. 2016. Richness, biomass, and nutrient content of a wetland macrophyte community affect soil nitrogen cycling in a diversity-ecosystem functioning experiment[J]. Ecological Engineering, 95: 252-265. DOI:10.1016/j.ecoleng.2016.06.057 |
Li Q S, Liu Y N, Du Y F, et al. 2011. The behavior of heavy metals in tidal flat sediments during fresh water leaching[J]. Chemosphere, 82(6): 834-838. DOI:10.1016/j.chemosphere.2010.11.026 |
Li X, Sun Z G, Tian L P, et al. 2020. Effects of spatial expansion between Phragmites australis and Cyperus malaccensis on variations of arsenic and heavy metals in decomposing litters in a typical subtropical estuary (Min River), China[J]. Chemosphere. |
吕国红, 周莉, 赵先丽, 等. 2006. 芦苇湿地土壤有机碳和全氮含量的垂直分布特征[J]. 应用生态学报, 17(3): 3384-3389. |
Nawaz M A, Jiao Y Y, Chen C, et al. 2018. Melatonin pretreatment improves vanadium stress tolerance of watermelon seedlings by reducing vanadium concentration in the leaves and regulating melatonin biosynthesis and antioxidant-related gene expression[J]. Journal of Plant Physiology, 220: 115127. |
聂兰仕. 2012.中国、欧洲与北美大陆地球化学元素分布特征及对比研究[D].北京: 中国地质大学. 1-157
|
秦先燕, 谢永宏, 陈心胜. 2010. 湿地植物间竞争和促进互作的研究进展[J]. 生态学杂志, 29(1): 117-123. |
Saco D, Martín S, San Jose P. 2013. Vanadium distribution in roots and leaves of Phaseolus vulgaris:morphological and ultrastructural effects[J]. Biologia Plantarum, 57(1): 128-132. DOI:10.1007/s10535-012-0133-z |
Sardans J, Peñuelas J. 2014. Climate and taxonomy underlie different elemental concentrations and stoichiometries of forest species:The optimum "biogeochemical niche"[J]. Plant Ecology, 215(4): 441-455. DOI:10.1007/s11258-014-0314-2 |
宋红丽, 孙志高, 牟晓杰, 等. 2012. 黄河三角洲新生湿地不同生境下翅碱蓬锰和锌含量的季节变化[J]. 湿地科学, 10(1): 65-73. |
Sun Z G, Li J B, He T, et al. 2019. Bioaccumulation of heavy metals by Cyperus malaccensis and Spartina alterniflora in a typical subtropical estuary (MinRiver) of Southeast China[J]. Journal of Soils and Sediments, 19: 2061-2075. DOI:10.1007/s11368-018-2177-y |
Sun Z G, Li J B, He T, et al. 2017. Spatial variation and toxicity assessment for heavy metals in sediments of intertidal zone in a typical subtropical estuary (Min River) of China[J]. Environmental Science & Pollution Research International, 24(29): 23080-23095. |
Sun Z G, Mou X J, Tong C, et al. 2015. Spatial variations and bioaccumulation of heavy metals in intertidal zone of the Yellow River estuary, China[J]. Catena, 126: 43-52. DOI:10.1016/j.catena.2014.10.037 |
Tong C, Zhang L H, Wang W Q, et al. 2011. Contrasting nutrient stocks and litter decomposition in stands of native and invasive species in a sub-tropical estuarine marsh[J]. Environmental Research, 111(7): 909-916. DOI:10.1016/j.envres.2011.05.023 |
谭海霞, 金照光, 孙富强, 等. 2019. 滦河口湿地植物-土壤生态化学计量相关性研究[J]. 水土保持研究, 26(2): 68-73. |
童晓雨, 孙志高, 曾阿莹, 等. 2019. 闽江河口互花米草海向入侵对湿地土壤无机硫赋存形态的影响[J]. 应用生态学报, 30(10): 3518-3526. |
王华, 孙志高, 李家兵, 等. 2019. 闽江口芦苇与短叶茳芏湿地土壤无机硫形态分布特征及其影响因素[J]. 生态学报, 39(13): 4921-4932. |
王为东, 王亮, 聂大刚, 等. 2010. 白洋淀芦苇型水陆交错带水化学动态及其净化功能研究[J]. 生态环境学报, 19(3): 537-543. |
王华, 孙志高, 李家兵, 等. 2018. 闽江口典型芦苇湿地与短叶茳芏湿地土壤碳氮含量的空间分布特征[J]. 生态学杂志, 37(4): 1102-1110. |
汪金舫, 刘铮. 1994. 钒在土壤中的含量分布和影响因素[J]. 土壤学报, 31(1): 61-67. |
王云, 魏复盛. 1995. 土壤环境元素化学[M]. 北京: 中国环境科学出版社.
|
Wan S Z, Yang G S, Mao R. 2020. Responses of leaf nitrogen and phosphorus allocation patterns to nutrient additions in a temperate freshwater wetland[J]. Ecological Indicators, 110: 105949. DOI:10.1016/j.ecolind.2019.105949 |
Yang J Y, Tang Y. 2015. Accumulation and biotransformation of vanadium in Opuntia microdasys[J]. Bulletin of Environmental Contamination and Toxicology, 94(4): 448-452. DOI:10.1007/s00128-015-1498-4 |
俞琳莺, 孙志高, 孙文广, 等. 2019. 外源氮输入对生长季黄河口碱蓬-土壤系统磷分布规律的影响[J]. 环境科学研究, 32(6): 1052-1062. |
祝贺, 孙志高, 衣华鹏, 等. 2016. 黄河口不同类型湿地土壤中钒和钴含量的空间分布特征[J]. 水土保持学报, 30(1): 315-320. |
曾英, 倪师军, 张成江. 2004. 钒的生物效应及其环境地球化学行为[J]. 地球科学进展, 19(S): 472-476. |
张晓晓. 2010.黄河下游水体及河口湿地沉积物中重金属的变化特征研究[D].青岛: 中国海洋大学. 1-80
|