 2020, Vol. 40
2020, Vol. 40



2. 生态环境部南京环境科学研究所, 南京 210042;
3. 中日友好环境保护中心, 北京 100029
2. Nanjing Institute of Environmental Science, Ministry of Ecology and Environment, Nanjing 210042;
3. Sino-Japan Friendship Center for Environmental Protection, Beijing 100029
随着滆湖富营养化问题的日益突出, 蓝藻“水华”、淡水资源缺少等问题日趋严重.淡水、浅水湖泊常与城市毗邻, 城市快速发展带来的面源或点源污染无法被全面控制, 加剧着湖泊的污染与富营养化, 湖泊生态平衡逐渐失稳.富营养化的直观表现包括频频暴发的蓝藻“水华”与变化明显的水环境因子, 而蓝藻“水华”的发生与作为水生态系统初级生产者的浮游植物之间息息相关(Ye et al., 2010; Magana-Arachchi et al., 2011; Jiang et al., 2017).
滆湖(119°48′E, 31°35′N)是太湖湖群第二大淡水湖泊, 位于太湖西北方向, 地处北亚热带季风气候区, 南北向纵跨常州市武进区与无锡宜兴市, 水域面积约166.7 km2, 常年水深约1.5 m, 每年5—9月是滆湖水华高发期, 大面积藻类覆盖湖面, 水体透明度大幅下降, 局部地区出现蓝藻堆积现象.目前研究太湖流域蓝藻的文章较多, 对滆湖研究较少, 并且一般关注滆湖北部的治理与修复(吴沛沛等, 2015; 何尚卫等, 2015; 高亚等, 2015; 陈志宁等, 2016; Xiong et al., 2016), 滆湖湖心区域乃至整个滆湖的整体性研究较少, 且缺乏对初期“水华”浮游植物群落变化特征及与水环境因子的相关性研究.本文对2018年5—7月的滆湖水环境因子、底泥相关指标以及浮游植物群落特征全面剖析, 初步探讨了浮游植物优势种群与水环境因子的相关性, 为揭示滆湖“水华”初期浮游植物群落变化特征及主导环境因子的内在联系提供参考, 以期为滆湖“水华”在发生初期得到有效遏制提供数据支撑.
2 材料与方法(Materials and methods) 2.1 研究区域及样品采集2018年5—7月, 蓝藻“水华”初期(谢小萍, 2016)在滆湖全湖采集水、底泥及浮游植物, 设如图 1的22个采样点.采集水面以下10 cm的柱状水样, YSI多参数水质检测仪现场测pH、溶解氧(DO)及水温(T).采两瓶水样, 每瓶1000 mL, 一瓶带回实验室立即加入10~15 mL的鲁哥试剂静置保存, 另一瓶水样过孔径0.7 μm的GF/F玻璃纤维滤膜后测试各项水质指标.加入鲁哥试剂后的水样倒入烧杯静置24~48 h, 虹吸法吸上覆水至量筒读数, 留下约10~20 mL底质保存待测.彼得森采泥器采集的底泥经烘干研磨过100目筛保存.
 |
| 图 1 采样点分布图 Fig. 1 Distribution map of Sampling points |
溶解性总磷(DTP)、溶解性总氮(DTN)与总有机碳(TOC):取50 mL原始水样经马弗炉450 ℃烘4 h的0.7 μm GF/F膜抽滤, 在50 mL比色管中稀释并加入与稀释水样1:1的碱性过硫酸钾作为氧化剂, 经高温灭菌锅121 ℃消解0.5 h后用钼锑抗分光光度法比色法分别在220 nm处测DTN, 700 nm处测DTP; 余下的过膜水样装入体积为24 mL的顶空瓶中, 液面高度在1/2到2/3之间, 使用总有机碳分析仪ASI-L(SHIMADZU CORPORATION, JAPAN)分析TOC.
水体悬浮物(SS)与悬浮物烧失量(G2):量筒计量为a(150~300 mL)的水样经马弗炉450 ℃烘过4 h的1.2 μm GF/C膜抽滤, 每张膜初始重量为m1, 滤过水样的膜以105 ℃烘箱烘4 h后重量为m2, 最终经450 ℃马弗炉烘4 h后重量为m3, SS计算公式为(m2-m1)/a×103, 单位为g·L-1, G2计算公式为(m2-m3)/m2 (国家环境保护总局《水和废水监测分析方法》委员会, 2002).
叶绿素a(Chl-a):马弗炉450 ℃烘过4 h的1.2 μm GF/C膜抽滤200 mL水样后, 将膜冷冻保存24 h, 然后使用分光光度法中的乙醇法进行测定(杨琳, 2019).
底泥有机质(OM)与有机磷(OP):OM使用重铬酸钾容量法——稀释热法检测, OP使用Ivanoff有机磷分级修正体系(郭旭欣, 2016)检测.
2.2.2 浮游植物多样性指数加入鲁哥试剂后吸至量筒中水体读数为N1, 保存待测的含底质水体含量记为N2, 移液枪取0.1 mL的底质到计数框(20 mm×20 mm), 立即盖上盖玻片(24 mm×24mm), 保证无气泡产生后在电子显微镜下以目镜行格法计数, 每瓶计数3片(国家环境保护总局《水和废水监测分析方法》委员会, 2002), 每片由ToupView软件拍摄50张照片后万深Algae C浮游生物计数检定系统进行计数, 浮游植物计数浓缩倍数按0.1N2/(N1+N2)计算.
2.3 数据处理与分析 2.3.1 多样性指数计算香农-威纳(Shannon-Wiener)指数H与均匀性(Pielou)指数J的描述:
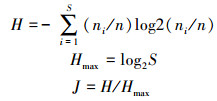
|
(1) |
式中, H为多样性指数, ni为样品中第i中生物的个数, n为样品所有种类的个数, S为样品的种类总数; J为均匀性指数.根据1990年沈韫芬等著写的《微型生物监测新技术》书中利用生物指数指示污染等级的划分, H>3为轻或无污染, H=1~3为中污染, H<1为重污染.J=0.5~0.8为轻污染, J=0.3~0.5为中污染, J=0~0.3为重污染(沈韫芬等, 1990).
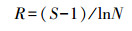
|
(2) |
式中, 物种丰富度(Margalef)指数R表示丰度, N为浮游植物总频数(曹方意等, 2014).R值大表明生态环境健康, 反之污染严重.
2.3.2 优势度计算优势度(Manaughton)指数Y的计算公式(3)为:
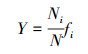
|
(3) |
式中, Ni为第i种藻类的数量, N为样品中藻类总数量, fi为第i种藻类出现的频率, 当Y>0.02时, 该种为优势种群(Lampitt et al., 1993).
2.3.3 相关性分析浮游植物群落结构与环境因子采用IBM SPSS Statistics 20进行Pearson相关性分析, 浮游植物优势种群藻类与环境因子的冗余分析(RDA)使用Canoco for Windows 5分析(Jindal et al., 2014; Zhang et al., 2018).
3 结果与分析(Results and analysis) 3.1 滆湖环境基本理化性质图 2a中5月份DTN浓度为0.17~2.2 mg·L-1, 均值为1.04 mg·L-1, 6月份DTN浓度为0.8~2.75 mg·L-1, 均值为1.53 mg·L-1, 7月份DTN浓度为0.56~2.88 mg·L-1, 均值为1.12 mg·L-1.5—7月DTN增加明显, 湖北区以北区域始终保持较高浓度并逐渐增长.滆湖南端的漕桥河口、北干河口等点位浓度较低, 且浓度较低点从5—7月呈范围性扩增, 浓度较高部分逐渐集中到湖心区域.DTP浓度(图 2b)大幅上涨, 5月份平均值为0.05 mg·L-1, 6月份上涨到0.1 mg·L-1, 7月份继续上涨至0.13 mg·L-1.5月份最高值为寨桥达到0.11 mg·L-1, 6月最高值在繁保区达到0.26 mg·L-1, 7月则在湖南区达到0.2 mg·L-1.其中繁保区除6月最高, 5月与7月含量均分别达到0.06 mg·L-1与0.21 mg·L-1.浓度较高点主要在湖心区域并从太滆运河向西南方向迁移(Tran et al., 2017), 在此期间, 揽月湾附近区域始终保持偏高浓度.图 2c与3d表明, pH较高值出现在万塔以及繁保区北附近, 5月份pH在8.03~9.26范围内, 均值为8.43, 各点相差较小, 6月升高到8.25~9.88, 均值为9.1, 初现差距, 7月则产生较大差距, pH范围7.68~10.75, 均值升高到9.43, 滆湖水华逐渐严重, 达到Gao等提到的9.0~10.5的范围(Gao et al., 2014), 此时pH较高值已向湖心区域迁移.5月DO为5.16~12.35 mg·L-1, 均值为7.35 mg·L-1, 6月DO为6.32~15.65 mg·L-1, 均值为10.08 mg·L-1, 7月DO为4.57~20.21 mg·L-1, 均值为10.21 mg·L-1, 滆湖湖北区以北部分各点DO均较低, 且随着时间推移递减, DO较高点在嘉泽往东方向以及往南沿岸方向, 且持续升高, 响应了pH的变化趋势, 且二者之间的响应关系符合Harris等的研究结果(Yen et al., 2004; Harris et al., 2016).
 |
| 图 2 5—7月各点位水质指标变化情况 (a.溶解性总氮, b.溶解性总磷, c.溶解氧, d.pH, e.烧失量, f.水体悬浮物, g.总有机碳, h.叶绿素a, i.水温, j.有机质) Fig. 2 Changes in water quality indicators at various sampling points from May to July (a.DTN, b.DTP, c.DO, d.pH, e.G2, f.SS, g.TOC, h.Chl-a, i.T, j.OM) |
 |
| 图 3 底泥各形态有机磷含量 (a.5月, b.6月, c.7月) Fig. 3 Contents of various forms of organic phosphorus in sediments (a. May, b.June, c. July) |
图 2f中SS迅速上升, 从5月份的0.03~0.08 mg·L-1上升到7月份的0.03~0.14 mg·L-1, 均值由0.05 mg·L-1变为0.09 mg·L-1, 上升约1倍.图 2e中烧失量G2由0.31%~3.62%上升到0.71%~7.97%, 5—6月的变化幅度不大, 6—7月骤然上升, 但相对于SS的上升, G2显然上升幅度较小(陈涛等, 2016).湖北区以北SS大幅上升, 但G2的上升幅度却小于其余点位, 表明湖北区以北各点水体悬浮物中含碳比例不高.同时, 总有机碳(TOC)数据(图 2g)表明, 5—7月湖北区以北区域TOC均值由7.11 mg·L-1下降为4.92 mg·L-1, 而剩下区域TOC平均含量由5.45 mg·L-1上升为6.40 mg·L-1, TOC含量的多少表明水体中存在相对应含量的有机污染物, TOC的变化率表明有机污染物逐渐由北向南至湖心区域迁移, 这与SS及G2分析结果一致.图 2h中叶绿素a(Chl-a)从均值3.93 mg·m-3上升到11.03 mg·m-3, 再大幅上涨到27.64 mg·m-3, 其中以湖心区域整体上涨幅度最大, 揽月湾、揽月湾西从低浓度骤然升高, 其余点为稳定中等浓度.
底泥中OM的变化如图 2j所示, 各点差异较大, 取水口的含量最少, 含量较高点主要集中在湖北区与湖南区, 达到30.324 g·kg-1与25.935 g·kg-1, 湖心区域与湖北区以北的OM含量偏高.滆湖底泥TN均值为2.97 mg·g-1, TP含量为1.12 mg·g-1, 图 3a~3c表明OP含量从多到少为钙结合态磷(HCl-P)、胡敏酸有机磷(Hum-P)、残渣态有机磷(Res-P)、弱吸附态磷(NaHCO3-P)、富里酸有机磷(Fulmic-P)(Zhao et al., 2019), 其中殷村港、漕桥河口以及湖北区与湖南区的OP总含量最高值分别达到266.97、227.89、265.51以及221.25 μg·g-1, 底泥中磷含量较高.
3.2 滆湖浮游植物时空分布特征 3.2.1 浮游植物群落结构分布特征浮游植物统计结果如图 4所示, 甲藻门为1%作为基数, 发现7个门, 单门占比超过5%的属有50个.5月占比最大为蓝藻门, 之后6—7月份绿藻门占比最大.优势属包括蓝藻门的鞘丝藻属与微囊藻属, 绿藻门的小球藻属与球团藻属, 隐藻门的回转隐藻属, 黄藻门的短圆柱单肠藻, 裸藻门的裸囊藻属, 5—7月, 蓝藻门的鞘丝藻属和微囊藻、绿藻门的小球藻属与球团藻逐渐占据主要优势, 绿藻门的新月藻属、栅藻逐渐增多.
 |
| 图 4 浮游植物分类占比 (a.5月, b.6月, c.7月) Fig. 4 The abundance Proportion of different phytoplankton (a. May, b.June, c. July) |
根据滆湖浮游植物优势种群分布特征(表 1), 将滆湖划为明显的3个分区, 分别是西部沿岸与滆湖南端区域, 鞘丝藻(A)与裸囊藻(M)占主要优势, 这部分区域以蓝藻门与裸藻门为主.然后是湖北区以北区域, 存在M或A但所占比例不高, 小球藻(C)、蛋白核小球藻(D)等绿藻门藻类占比增大, 其中揽月湾及揽月湾西等北部沿岸点具有明显的蓝藻堆积现象, 微囊藻(B)比例大幅上升.最后是湖心区域, 这块区域的主要浮游植物与湖北区以北区域类似, 但C、D的比例上升, 并出现新月藻(L)、斜生栅藻(I)等藻类.3块区域的种群个数也存在差异, 湖心区域的湖南区、北干河口、繁保区、寨桥及太滆河口种群个数偏低, 其余各点的种群个数以此为分界往南为递增趋势, 往北至大洪港亦为递增趋势.
| 表 1 浮游植物优势种群分布 Table 1 Phytoplankton dominant population distribution |
Shannon-Wiener指数H从2.6342~3.8922降低到2.2091~3.1924, 平均值从3.2928降到2.7811, 根据H值的污染等级划分, 滆湖整体由轻污染上升到中污染, 湖心区域各点与揽月湾及揽月湾西的多样性指数小于平均值均为中污染.Pielou指数J均值由0.6881下降到0.5963, 变化较小, 且根据J值的污染等级划分表现为轻污染, 其中揽月湾、揽月湾西、太滆河口、繁保区北、繁保区、湖南区以及寨桥在6—7月的J指数达到中污染, 与H指数的均值较低点响应.3块分区的每月各点间均匀性指数差异不大, 说明同一分区内浮游植物分布较均匀, 物种奇异度低, 可能是由于主要优势藻类占比逐渐增大, 如湖北区以北区域蓝藻门的鞘丝藻, 湖心区域绿藻门的小球藻占比增加(苏玉等, 2011; 李娣等, 2014).Margalef指数R均值从4.5278下降到3.9828, 其中最小值为繁保区北从2.9832下降到2.6681, 太滆河口、湖南区及繁保区等属于丰度较低的集中区域, 揽月湾与揽月湾西也低于均值, 根据丰度越低污染越严重的原则, 这两处区域属于滆湖中污染偏重区域, 且在6—7月间达到峰值.
| 表 2 浮游植物生物指数 Table 2 Phytoplankton Biology Index |
滆湖蓝藻“水华”除水体表面透明度的直观反馈, 还可以通过浮游植物群落变化特征, 以及叶绿素a(Chl-a)的变化反应滆湖蓝藻“水华”.本文为了解水、底泥指标或浮游植物对“水华”的影响, 对Chl-a与水质指标、浮游优势种群与环境因子、底泥OM与OP等进行相关性分析(Sondergaard et al., 2011), 以期得到“水华”初期的主导环境因素.
从图 5a、5b、5e中发现T与多项指标有相关性, 与pH、DO及Chl-a在0.01水平(双侧)显著相关, 且Pearson相关性分别为0.749、0.581与0.58, 均为中度以上相关性.表明T在水华发生过程中为主导因素之一(Sylwia et al., 2019).
 |
| 图 5 环境因子之间的相关性 Fig. 5 Correlation between environmental factors |
图 5c是5—7月Chl-a与DTP的相关性分析, 全程0.01水平(双侧)显著相关, 且Pearson相关性达到0.672呈中高度相关性, 表明DTP的变化很好地响应了叶绿素a的升降(董云仙等, 2011), 表明与蓝藻暴发存在紧密联系(Li et al., 2012).
图 5d是底泥OM与OP的相关性分析, 两者之间呈显著相关性(p < 0.05), 且Pearson相关性系数为0.892, 为高度相关关系, 表明蓝藻暴发期间, OM与OP同步释放, 底泥OP的释放与OM紧密相关, 主要释放出弱吸附态磷(NaHCO3-P)与富里酸有机磷(Fulmic-P), 在蓝藻“水华”较严重区如湖南区附近的湖心区域OM与OP均为高含量状态.
Chl-a在文献中常被直接用以表达蓝藻“水华”严重程度(邢建伟等, 2014), 当浮游植物大量繁殖, SS含量明显上升, Chl-a与SS的相关性(图 5f)表明两者之间p=0.006明显小于0.05, 在0.01水平(双侧)上显著相关.同时, 滆湖水体自然稳定状态下的浮游植物丰富度指数与Chl-a之间理论上呈一定相关性, 但由于蓝藻“水华”初期浮游植物种群更替与迁移以及优势种群占比的增加, 滆湖的Chl-a与滆湖浮游植物的丰富度指数呈不全面的相关, 响应较明显的有场北河口(R指数2.6376)、万塔(R指数2.878)及大洪港(R指数3.5849)等丰富度指数较低的点, 3个对应的Chl-a含量也均较低, 从5—7月分别为0.84~7.81、3.07~10.18、0.98~5.72 mg·m-3, 这与杜胜蓝等(2011)的研究结果一致.
由滆湖5—7月浮游植物优势种与环境因子的RDA排序图(夏莹霏等, 2019)(图 6)以及相关关系(表 3)可见, 鞘丝藻属与G2、DO、pH呈显著相关性, 与TOC呈负相关; 微囊藻属与DTP、OM、DTN呈显著相关, 与T呈负相关; 小球藻属与DTN呈显著相关, 与DTP、DO、pH、T呈负相关; 球团藻属与T呈显著相关, 新月藻属与T呈显著相关; 栅藻属、裸囊藻属与DTP呈显著相关; 回转隐藻属与OM、T显著相关; 短圆柱单肠藻与DTN呈显著相关, 与G2、pH、DO呈负相关.
 |
| 图 6 浮游植物优势种与环境因子的RDA排序图 (a为鞘丝藻属, b为微囊藻属, c为小球藻属, d为球团藻属, e为栅藻属, f为新月藻属, g为裸囊藻属, h为隐藻属, i为单肠藻属单) Fig. 6 RDA biplot of dominant species and environmental factors of phytoplankton |
| 表 3 滆湖环境因子与浮游植物优势种的相关关系 Table 3 Correlation between environmental factors and phytoplankton in Gehu Lake |
水质检测结果表明, 滆湖存在明显湖区分布差异, 湖北区以北沿岸各点水质状况最差, 王彧对滆湖水质富营养化调查中也表明滆湖北部污染较严重(王彧, 2016), 且揽月湾附近出现明显的蓝藻堆积(赵汉取等, 2009; Zhang et al., 2013; Wu et al., 2017), 阻碍自然光照, 使沉水植物生长受到抑制(尚媛媛等, 2015), 底泥OM含量逐渐升高且大于上覆水有机物含量增加比例, 响应了G2、TOC及底泥OM的变化趋势; 湖心区域劣等水质范围最广, Chl-a达到全湖最高, 上覆水有机物含量逐渐升高且范围逐渐扩大, 5—7月水质较差区域逐渐向滆湖湖心区域集中, DTP浓度最高点出现在湖心区域的繁保, 这可能与存在的养殖围网有关(胡莉莉等, 1991); 水质较好的区域主要是西部沿岸及滆湖南端湖区.T的变化趋势与水质因子在空间上的分布及变化趋势相关联(蔡金傍等, 2018), 尤其与DTP、pH、DO相关性较高, 从5—7月T的折线图变化趋势可以看出湖心区域与湖北区以北各点的T明显升高, 从26.4 ℃上升到32.6 ℃, 滆湖“水华”期T的变化特征与宗梅等总结的巢湖万年埠26~32 ℃的变化范围相近(宗梅等, 2016), 王华等对滇池的研究中表明T在7月份达到峰值后与浮游植物响应明显(王华等, 2016), 表明T是浅水湖泊“水华”发生互通的主导因素, 不仅仅对滆湖影响较大.湖心区域与湖北区以北区域OP与OM含量较高, 此外, 滆湖南部的殷村港与漕桥河口底泥OP含量高可能是由于所处范围的沉水植物较多导致, 底泥OM与OP的变化表明应对滆湖“水华”, 上覆水与底泥中的磷需同步关注(Cao et al., 2016).
4.2 不同湖区浮游植物分布特征湖心区域与湖北区以北区域多样性指数小于平均值, Shannon-Wiener指数H较高点位主要在滆湖南端区域, 多样性降低说明存在浮游优势种群泛滥.Pielou指数J划分的中污染区域主要为湖心区域与揽月湾及揽月湾西, 与H指数响应, 但整体上J指数表现为轻污染, 与H指数相关性不明显, 但综合分析表明5—7月湖心与北部沿岸区域富营养化程度较高, 整体表现为中污染(李清雪等, 1999), 且该两处区域Margalef指数R在滆湖中属于丰度较低区域.总体而言, 湖心区域与北部沿岸区域为“水华”严重区, 该区域浮游植物特征为种群个数较少, 多样性与丰度低, 且具有较明显的优势种群(陈洋, 2017)小球藻与球团藻, 同时栅藻随时间推移逐渐增多, 朱伟等做出的研究表明这与温度的提升有关(朱伟等, 2008).除两个“水华”严重区域, 西部沿岸及滆湖南端的浮游植物主要是鞘丝藻及裸囊藻为主, 对水华发生的贡献不大.5—7月, 滆湖绿藻门逐渐占据优势, 蓝藻门紧随其后, 形成“绿藻+蓝藻”型湖泊, 这与巢湖流域和太湖的浮游植物群落结构特征相似, 但巢湖与太湖中硅藻门也是优势门之一(路娜等, 2010; 叶琳琳等, 2017; 邱伟建等, 2018), 而滆湖蓝藻严重区域主要是绿藻门的小球藻与球团藻及蓝藻门的微囊藻作贡献, 这与曹方意等总结的“蓝藻+绿藻”型滆湖结果相同(曹方意等, 2014).
4.3 浮游植物群落与环境因子相关性环境因子差异与浮游植物优势种的分布特征将滆湖分成3部分, 按浮游植物现存量从少到多主要分为西部沿岸与滆湖南端区域、湖心区域及湖北区以北区域.湖心区域与湖北区以北区域的浮游优势种差异较小(陈立靖等, 2008), 以蓝藻门的鞘丝藻、微囊藻, 绿藻门的小球藻、球团藻为主, RDA分析显示与T、pH、DO、DTP及OM呈显著相关, 湖心区域渐增的新月藻属与栅藻分别与T和DTP相关, OP与OM的Pearson相关分析呈强相关, 表明两处“水华”严重区的变化与T相关明显(Feng et al., 2016), 且该变化不仅与水体的磷有关, 同样受底泥中磷的影响, 陈宇炜等在对太湖浮游植物的长期演变研究中表明微囊藻的增多与T和DTP也密切相关(Chen et al., 2003).西部沿岸与滆湖南端区域优势种为裸囊藻与鞘丝藻, 与TOC、T、pH及DTP相关, 由于滆湖南端的殷村港附近属于草型湖区(丁轶睿等, 2017), 植物自然衰亡带来TOC与磷素上升, 但整体水质依旧优于湖心区域及湖北区以北区域.Chl-a与OM及浮游植物种群密度不完全相关, 部分OM或浮游植物种群密度较高点位的Chl-a含量较低, 这可能与Kim等提出的OM及浮游植物种群密度增加到一定量后抑制底栖蓝藻的生长有关(Kim et al., 2016).总体而言, 滆湖水华初期藻类的主要相关因子包括T、pH、DO及水体与底泥中的磷, 朱文昌对梅梁湖浮游植物的相关环境因子研究同样表明总磷、总氮与水温为影响浮游植物生长的主要因素, 他还提出风速也是关键因素(朱文昌等, 2010), 高子涵对大通湖的RDA分析中也提出pH与T以及总磷是影响浮游植物群落结构的第一排序轴的重要影响因子(高子涵等, 2016).滆湖5—7月的pH基本呈弱碱性, 因此提高了浮游植物对CO2吸收的能力, 从而提高生产力(Zhang et al., 2011), T从26.1 ℃快速上升后基本维持在29.2~32.4 ℃, T不仅与浮游植物间存在相关, 与pH、DO、Chl-a等多项环境因子彼此也显著相关(RosiÅ et al., 2017).根据相关性结果, 需对T、pH及DO作预警性监测, 同时关注水体及底泥中的磷, 预先对磷采取钝化及去除措施(秦伯强等, 2007).
5 结论(Conclusions)1) 滆湖“水华”初期暴发点发生在6—7月, 湖心区域最严重, T、Chl-a与DTP含量较高并呈上升趋势.湖北区以北的揽月湾等沿岸各点存在明显的蓝藻堆积现象, 底泥OM与OP, 水体SS与TOC含量彼此间的响应表明湖北区以北区域有机物含量较高并大量沉积于底泥.
2) Shannon-Wiener指数H表明, 由于湖心区域及揽月湾附近点位优势种群占比过大形成泛滥使浮游植物多样性降低, Pielou指数J下降到0.3~0.5间, 整体由轻污染变为中污染, 污染程度提升, 其中主要浮游植物优势种为蓝藻门的鞘丝藻与微囊藻以及绿藻门的小球藻, “水华”严重期(6—7月)浮游植物主要是绿藻门占比最高, 其次为蓝藻门.
3) 滆湖5—7月的Chl-a与DTP呈高度相关, 与SS呈中度相关; 水温T与DO、pH、Chl-a呈中高度相关; RDA排序图表明“水华”初期的藻类优势种主要与T、DTP、OP、DO以及pH相关, 其中T与DTP为主导环境因素, T达到26~30 ℃时应采取预防措施, 对初期“水华”的控制不仅需要关注水中的磷, 底泥中磷的释放应同样重点关注.
Baek J S, Kim Y J, Youn S J. 2016. Cyanobacteria community and growth potential test in sediment of Lake Paldang[J]. Journal of Korean Society on Water Environment, 32(3): 261-270. DOI:10.15681/KSWE.2016.32.3.261 |
Cao X, He J, Wang Y, et al. 2016. Phosphorus mobility among sediments, water and cyanobacteria enhanced by cyanobacteria blooms in eutrophic Lake Dianchi(Article)[J]. Environmental Pollution, 219: 580-587. DOI:10.1016/j.envpol.2016.06.017 |
Chao J Y, Duan J C, Zhang Y M, et al.2013.Study on Application of Pre-Dam in the Eutrophication Control of Gehu Lake Lake[M].PROCEEDINGS of the 35TH IAHR WORLD CONGRESS, VOLS I and II.3158-3165
|
Chen Y W, Qin B Q, Teubner K, et al. 2003. Long-term dynamics of phytoplankton assemblages:Microcystis domination in Lake Taihu, a large shallow lake in China[J]. Journal of Plankton Reasearch, 25(1): 445-453. |
Cieszyńska A, Konik M, Śliwińska-Wilczewska S, et al. 2019. Environmental drivers of bloom-forming cyanobacteria in the Baltic Sea:Effects of salinity, temperature, and irradiance[J]. Estuarine Coastal & Shelf Science: 139-150. |
蔡金傍, 孙旭, 苏良湖, 等. 2018. 滆湖污染源调查与分析[J]. 江苏农业科学, 46(5): 224-227. |
曹方意, 龙建, 夏霆. 2014. 滆湖浮游藻类分布特性及水质评价[J]. 西南给排水, 36(5): 49-53. |
陈立婧, 彭自然, 孔优佳, 等. 2008. 江苏滆湖浮游藻类群落结构特征[J]. 生态学杂志, 27(9): 1549-1556. |
陈涛, 杨波, 张际标, 等. 2016. 湛江湾表层海水悬浮物分布特征及影响因素[J]. 环境科学与技术, 39(S2): 422-428. |
陈洋. 2017. 太湖不同区域浮游植物群落结构特征及其与环境因子的关系[J]. 水生态学杂志, 38(3): 38-44. |
陈志宁, 张红高, 周崴, 等. 2016. 滆湖大型底栖动物群落分布和氮磷因子的相关分析[J]. 环境监控与预警, 8(1): 45-50, 59. DOI:10.3969/j.issn.1674-6732.2016.01.015 |
邓建才, 路娜, 尹洪斌, 等. 2010. 巢湖流域春季浮游植物群落结构特征及其与环境因子的关系[J]. 湖泊科学, 22(6): 950-956. |
丁轶睿, 李定龙, 张毅敏, 等. 2017. 滆湖底泥细菌群落结构及多样性[J]. 环境科学学报, 37(5): 1649-1656. |
董云仙, 洪雪花, 谭志卫, 等. 2012. 高原深水湖泊程海氮磷形态分布特征及其与叶绿素a的相关性[J]. 生态环境学报, 21(2): 333-337. DOI:10.3969/j.issn.1674-5906.2012.02.023 |
杜胜蓝, 黄岁樑, 臧常娟, 等. 2011. 浮游植物现存量表征指标间相关性研究Ⅰ:叶绿素a与生物量[J]. 水资源与水工程学报, 22(1): 40-44. |
Diane K S, Jeffrey C C, Yong H G, et al. 2014. Influence of cyanobacteria blooms on sediment biogeochemistry and nutrient fluxes[J]. Limnology and Oceanography, 59(3): 959-971. DOI:10.4319/lo.2014.59.3.0959 |
Dondajewska R, Kozak A, RosiÅska J, et al. 2017. Cyanobacteria blooms before and during the restoration process of a shallow urban lake[J]. Journal of Environmental Management, 198(Pt 1): 340-347. |
Feng L J, Liu S Y, Wu W X, et al. 2016. Dominant genera of cyanobacteria in Lake Taihu and their relationships with environmental factors[J]. Journal of Microbiology, 54(7): 468-476. DOI:10.1007/s12275-016-6037-4 |
房晟忠, 王华, 杨树平, 等. 2016. 滇池浮游植物群落特征及与环境因子的典范对应分析[J]. 中国环境科学, 36(2): 544-552. DOI:10.3969/j.issn.1000-6923.2016.02.034 |
高亚, 李勇, 潘继征, 等. 2015. 江苏滆湖北部区整治后浮游植物时空分布及环境因子变化规律[J]. 湖泊科学, 27(4): 649-656. |
高子涵, 孟宪智, 周绪申. 2016. 大通湖浮游植物群落结构与环境因子关系[J]. 水利信息化, (5): 21-30. |
关保华, 尚媛媛, 郑建伟, 等. 2015. 蓝藻堆积对水环境和沉水植物生长的影响[J]. 中国农学通报, 31(5): 195-198. |
郭旭欣. 2016. 容量法测定土壤有机质2种加热方法比较研究[J]. 现代农业科技, (9): 206-209. DOI:10.3969/j.issn.1007-5739.2016.09.124 |
国家环境保护总局《水和废水监测分析方法》编委会. 2002. 水和废水监测分析方法:第4版[M]. 北京: 中国环境科学出版社.
|
Harris J M, Kularatne R K A, Vinobaba P, et al. 2016. Spatial and temporal distribution of cyanobacteria in Batticaloa Lagoon[J]. Journal of Environmental Sciences, 47: 211-218. DOI:10.1016/j.jes.2016.01.020 |
Huang T C, Yen U C, Yen T C. 2004. Observation of the circadian photosynthetic rhythm in cyanobacteria with a dissolved-oxygen meter[J]. Plant Science, 166(4): 949-952. DOI:10.1016/j.plantsci.2003.12.005 |
杭鑫, 李亚春, 谢小萍, 等. 2016. 气温对太湖蓝藻复苏和休眠进程的影响[J]. 湖泊科学, 28(4): 818-824. |
何尚卫, 李勇, 潘继征, 等. 2015. 江苏滆湖北部湖区综合整治后夏季水体光环境研究[J]. 湖泊科学, 27(2): 311-318. |
胡莉莉, 赵瑞坤, 张向群, 等. 1991. 滆湖网围养殖后对水体富营养化的影响[J]. 水产学报, 15(4): 291-301. |
胡晓东, 刘劲松, 吴沛沛, 等. 2015. 滆湖北部底泥疏浚的生态效应研究[J]. 水生态学杂志, 36(2): 32-38. |
胡晓东, 夏莹霏, 徐季雄, 等. 2019. 太湖浮游植物功能群季节演替特征及水质评价[J]. 湖泊科学, 31(1): 134-146. |
黄晓峰, 邱伟建, 钱程远, 等. 2018. 莲花荡浮游植物群落结构与环境因子的相关性[J]. 环境科学与技术, 41(S2): 1-8. |
Jiang J, Li Y C, Wang N, Zhang, et al. 2017. Effect of cyanobacteria aggregation on the migration and release of heavy metals in Lake Chaohu[J]. Hupo Kexue/Journal of Lake Sciences, 29(3): 558-566. DOI:10.18307/2017.0304 |
Jiang M, Sheng Y Q, Zhao G Q, et al. 2019. The biogeochemical characteristics of phosphorus in coastal sediments under high salinity and dredging conditions[J]. Chemosphere: 681-692. |
Jindal R, Singh U B, Thakur R K, et al. 2014. Phytoplankton dynamics and species diversity in a shallow eutrophic, natural mid-altitude lake in Himachal Pradesh (India):Role of physicochemical factors[J]. Chemistry and Ecology, 30(4): 328-338. |
Lampitt R S, Turley C M F, Wishner K. 1993. Marine snow studies in the Northeast Atlantic Ocean:distribution, composition and role as a food source for migrating plankton[J]. Marine Biology, 116(4): 689-702. DOI:10.1007/BF00355486 |
Larsen S E, Jorgensen T B, Sondergaard M, et al. 2011. Using chlorophyll a and cyanobacteria in the ecological classification of lakes[J]. Ecological Indicators, 11(5): 1403-1412. DOI:10.1016/j.ecolind.2011.03.002 |
Li W C, Pan J Z, Wu X D, et al. 2017. Restoration in northern Lake Gehu Lake, a eutrophic lake in China[J]. Chinese Journal of Oceanology and Limnology, 35(6): 1417-1431. DOI:10.1007/s00343-017-6107-4 |
李娣, 李旭文, 牛志春, 等. 2014. 太湖浮游植物群落结构及其与水质指标间的关系[J]. 生态环境学报, 23(11): 1814-1820. DOI:10.3969/j.issn.1674-5906.2014.11.014 |
李清雪, 陶建华. 1999. 应用浮游植物群落结构指数评价海域富营养化[J]. 中国环境科学, 19(6): 548-551. DOI:10.3321/j.issn:1000-6923.1999.06.017 |
刘波, 吴晓东, 叶琳琳, 等. 2017. 太湖西北湖区浮游植物和无机、有机氮的时空分布特征[J]. 湖泊科学, 29(4): 859-869. |
陆敏, 石浚哲, 朱文昌. 2010. 梅梁湖水体浮游植物与环境因子的关系[J]. 环境监测管理与技术, 22(3): 27-30. DOI:10.3969/j.issn.1006-2009.2010.03.007 |
Li Y, Li D. 2012. Competition between toxic Microcystis aeruginosa and nontoxic Microcystis wesenbergii with Anabaena PCC7120[J]. Journal of Applied Phycology, 24(1): 69-78. DOI:10.1007/s10811-010-9648-x |
Magana-Arachchi D, Liyanage M, Wanigatunge R. 2011. Molecular characterization of cyanobacterial diversity in Lake Gregory, Sri Lanka[J]. Chinese Journal of Oceanology and Limnology, 29(4): 898-904. DOI:10.1007/s00343-011-0519-3 |
秦伯强, 汤祥明, 王小冬, 等. 2007. 太湖富营养化与蓝藻水华引起的饮用水危机-原因与对策[J]. 地球科学进展, 22(9): 896-906. DOI:10.3321/j.issn:1001-8166.2007.09.003 |
Sheng Y Q, Wang J, Zhao G Q, et al. 2018. Optimized digestion methods:organic phosphorus sequential extraction, total phosphorus, and nitrogen simultaneous determination in sediments[J]. Journal of Soils and Sediments, 18(5): 2072-2080. DOI:10.1007/s11368-018-1959-6 |
沈韫芬, 顾曼如, 龚循矩, 等. 1990. 微型生物监测新技术[M]. 北京: 中国建筑工业出版社.
|
绳秀珍, 邢建伟, 线薇微. 2014. 2012年夏季长江口颗粒有机碳、氮分布特征及其来源[J]. 环境科学, 35(7): 2520-2527. |
苏玉, 文航, 王东伟, 等. 2011. 太湖武进港区域浮游植物群落特征及其主要水质污染影响因子分析[J]. 环境科学, 32(7): 1945-1951. |
Tan X, Wu X D, Ye L L, et al. 2010. Cell Lysis of Cyanobacteria and Its Implications for Nutrient Dynamics[J]. International Review of Hydrobiology, 95(3): 235-245. DOI:10.1002/iroh.200911194 |
Tran N B, Tang V T, Tang T T. 2017. 太滆运河污染源调查与分析[J]. 安徽农学通报, 23(20): 66-68. DOI:10.3969/j.issn.1007-7731.2017.20.029 |
Wu X D, Xiong C H, Zhang R L, et al. 2016. Distribution and pollution assessment of nutrient and heavy metals in surface sediments from lake Gehu Lake in Southern Jiangsu Province, China[J]. Journal of Environmental Sciences, 37(3): 925-934. |
万蕾, 朱伟, 赵联芳. 2008. 不同温度和营养盐质量浓度条件下藻类的种间竞争规律[J]. 生态环境学报, 17(1): 6-11. DOI:10.3969/j.issn.1674-5906.2008.01.002 |
王翔, 宇宗梅, 郑西强. 2016. 巢湖万年埠沿岸浮游植物与环境因子关系[J]. 安徽农业科学, 44(27): 102-105. DOI:10.3969/j.issn.0517-6611.2016.27.035 |
王彧. 2016. 滆湖水质富营养化调查与评价[J]. 淮海工学院学报(自然科学版), 25(3): 88-91. DOI:10.3969/j.issn.1672-6685.2016.03.019 |
韦肖杭, 姚伟忠, 赵汉取, 等. 2009. 蓝藻暴发后南太湖水域浮游生物及富营养化[J]. 浙江海洋学院学报(自然科学版), 28(1): 21-24, 35. DOI:10.3969/j.issn.1008-830X.2009.01.004 |
Xu P, Zhang S H, Zhang K X, et al. 2018. Phosphorus release from cyanobacterial blooms during their decline period in eutrophic Dianchi Lake, China[J]. Environmental Science & Pollution Research, 25(14): 13579-13588. |
杨琳. 2019. 浮游植物叶绿素a测定方法研究进展[J]. 四川环境, 38(1): 156-160. |
Zhang Q T, Hu G K. 2011. Effect of nitrogen to phosphorus ratios on cell proliferation in marine micro algae[J]. Chinese Journal of Oceanology and Limnology, 29(4): 739-745. DOI:10.1007/s00343-011-0503-y |