 2020, Vol. 40
2020, Vol. 40



2. 哈尔滨师范大学地理科学学院, 哈尔滨 150025
2. College of Geographical Science, Harbin Normal University, Harbin 150025
全球气候变暖已成为全世界共同关注的环境问题(Mearns et al., 1984).研究证明, 近几十年来气温升高主要是温室气体排放造成的, 而CO2、CH4和N2O被认为是最重要的温室气体(IPCC, 2007).其中CO2是全球最重要的温室气体, 与CO2相比, CH4增温潜势大约是CO2的23倍, 是仅次于CO2的全球第二大温室气体, 对全球气候变暖贡献率为14%(IPCC, 2007).N2O的增温效应约是CO2的150~200倍(Grosso et al., 2005), 而大气中N2O对温室气体贡献率为8%(IPCC, 2007).在过去的几十年里, CO2、CH4和N2O以年均0.5%、0.8%和0.3%的速度在大气中迅速增长.
凋落物是生态系统分解者亚系统的重要组成部分, 是土壤碳、氮的重要来源.凋落物在维持土壤肥力、促进生态系统正常的物质循环和养分平衡方面有着重要的作用(Sayer et al., 2006; Sayer et al., 2011).凋落物本身也会通过与土壤水热因子, 土壤微环境以及土壤生物的相互作用, 直接或间接地影响土壤温室气体(CO2、CH4、N2O)排放(Niu et al., 2009;高强等, 2016; Sangsub et al., 2017;张素彦等, 2018).凋落物是地表土壤微生物生长代谢的主要底物来源之一, 为其提供生长代谢所需物质及能量, 并在分解过程中释放CO2, 因此凋落物质量与数量的变化不可避免地引起土壤呼吸的变化.凋落物层也可通过改变小气候和土壤理化性质来影响碳循环, 从而对土壤CH4排放产生重要的影响.湿地是陆地上重要的生态系统, 虽然仅占地球表面积的5%~8%, 却储存了全球20%~30%的碳.三江平原作为东北地区乃至全国最大的平原湿地, 其温室气体“源”和“汇”的角色转变对区域、全国乃至全球碳氮循环具有重要影响.目前, 对于凋落物的研究主要集中在森林以及草地生态系统中, 并且主要研究凋落物分解及其影响机制, 而对于凋落物处理下温室气体如何响应相对研究较少, 尤其是对于湿地生态系统中凋落物对温室气体排放的贡献如何还需要进一步研究去揭示.
为此, 本研究采取野外原位控制实验, 分析不同凋落物处理对三江平原小叶章湿地土壤温度和含水量的影响, 明确不同凋落物处理下小叶章湿地温室气体排放的主要驱动因子.既可以为揭示湿地生态系统温室气体排放对凋落物的响应机制提供理论依据, 也可以为未来全球变化中湿地生态系统的科学管理以及生态保护提供有价值的参考.
2 研究地区与研究方法(Research area and methods) 2.1 研究区概况研究地点设置在黑龙江省科学院自然与生态研究所三江平原湿地生态定位研究站内(图 1).研究区海拔为55~65 m, 属温带湿润半湿润季风气候, 最热月为7月, 最冷月为1月, 年平均气温为1.9 ℃.多年平均降水量为585 mm.植被类型以小叶章(Calamagrostis angustifolia)为建群种, 主要伴生种包括漂筏苔草(Carex pseudocuraica)、灰脉苔草(Carex appendiculata)、狭叶甜茅(Glyceria spiculosa)以及毛果苔草(Carex lasiocarpa)等.样地土壤类型主要由草甸土和白浆土组成.具体的土壤理化性状见表 1.
 |
| 图 1 实验区位置-洪河国家级自然保护区 Fig. 1 Location of sampling site in the Sanjiang Plain-Honghe national nature reserve |
| 表 1 土壤基本理化性质 Table 1 The basic physical and chemical properties of soil |
利用黑龙江省科学院自然与生态研究所三江平原湿地生态定位研究站内的试验平台.每个处理面积为1 m×1 m, 处理1:去除当年凋落物和草碳层(NL+NH, 土壤裸露);处理2:仅去除当年凋落物(NL);处理3:自然状态(对照, CK);处理4:倍增当年凋落物(DL);处理5:倍增当年凋落物和倍增草碳层(DL+DH).对于处理1:第一次处理时, 首先将当年凋落物和草碳层移走, 裸露土壤, 此后收集样地内凋落物, 防止凋落物进入样地;处理2:仅将当年凋落物移走, 此后收集样地内凋落物, 防止凋落物进入样地;处理3:凋落物自然状态, 允许凋落物正常进入;处理4:首先将处理2去除的当年凋落物, 平均撒在处理4的样地内, 此后每月将处理2收集的凋落物等面积均匀撒在处理4样地内;处理5:首先把样地当年凋落物取出, 然后把处理1去除的草碳层均匀覆盖到处理5样地原有草碳层上, 然后把处理5样地之前取出的当年凋落物和处理1去除的当年凋落物平均撒在处理5的样地内, 此后, 每个月将处理1收集的凋落物等面积均匀撒在处理5样地内.每个处理设置3个重复.
2.3 气体样品采集与分析CO2、CH4和N2O 3种温室气体采集利用静态箱法, 气体采样箱是由不锈钢材料制作, 长50 cm、宽50 cm、高50 cm.并在每个静态箱下面安装一个不锈钢底座, 底座长50 cm、宽50 cm、高20 cm, 并将其插入土壤中20 cm.每次气体样品采集时, 为了确保采样时的密闭效果, 防止空气中气体进入采样箱内, 首先在底座的凹槽内注满水, 然后在将采样箱安放在凹槽上.为了减少外界温度对采样箱内气体交换造成影响, 每个采集箱外部都用保温板进行处理.使用100 mL注射器采集气体样品, 每间隔10 min采集一次样品(每个处理采集4次).试验周期为2017.05—2017.09, 每7 d采集一次气体, 如遇极端天气延后1 d采集, 每次取样时间为上午9:00—11:00.在气体样品取样的同时, 同步测定土壤温湿度, 并且记录采样箱内温度以及采样箱外大气温度.采集的气体样品使用Agilent7890B气相色谱仪进行浓度分析(秦晓波等, 2006;张艺等, 2016).
2.4 气体通量计算公式
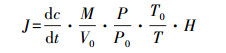
|
(1) |
式中, J为气体通量(mg·m-2·h-1);M为测定气体的摩尔质量;c为气体浓度(μg·m-3);dc/dt为采样时气体浓度随着采样时间变化的直线斜率;H为采样箱内气体高度(地表无积水时为静态箱高度, 地表有积水时为静态箱高度减去积水深度);P为采样点气压;T为气体采集时的绝对温度;V0、P0、T0分别为标准状态下的体积、空气绝对温度和气压(Wickland et al., 2001).
2.5 土壤温湿度测定土壤温度和含水量采用5TE土壤温湿度仪监测(每隔10 min记录1次).并于每次气体取样时同时测定土壤10 cm深度土壤温湿度.
2.6 数据处理采用Excel 2016和SPSS 22.0软件对数据进行统计分析.采用单因素(one-way ANOVA)和Duncan法进行方差分析和多重比较(α=0.05), 用回归分析法对土壤温湿度和温室气体排放通量进行相关分析.作图采用SigmaPlot软件.图表中数据代表平均值±标准差.
3 结果与分析(Results and analysis) 3.1 不同凋落物处理下10 cm土壤温度的时间动态从整个测定时期来看可以分为3个阶段(图 2), 春季阶段(2017年5月27日—2017年6月21日)温度为7.6~14.1 ℃, 夏季阶段(2017年7月2日—2014年8月23日)温度为15.3~21.2 ℃, 秋季阶段(2017年9月1日—2017年10月5日)温度为13.4~18.6 ℃.5个凋落物处理样地土壤温度在时间上变化基本一致, 不同凋落物处理样地均出现先升高后降低的趋势(图 2).5个样地的土壤温度在5月27日时都较低.在进入6月份之后, 开始逐渐上升, 去除当年凋落物和草炭层样地在7月14日达到最大值(21.2 ℃).整个生长季中, 去除当年凋落物和草炭层样地的土壤温度始终高于其它处理.进入9月份, 地温迅速下降, 5个处理的温度逐渐趋于一致(图 3).
 |
| 图 2 小叶章湿地土壤温度季节动态 Fig. 2 Seasonal variations of soil temperature in Calamagrostis angustifolia wetland |
 |
| 图 3 不同凋落物处理下小叶章湿地土壤温度 Fig. 3 Soil temperature of Calamagrostis angustifolia wetland underdifferent litter treatment |
5个凋落物处理样地土壤含水量在时间上变化基本一致, 不同凋落物处理样地均出现先降低后升高然后缓慢的趋于平稳的趋势(图 4).在生长季初期, 由于冻融期刚刚结束, 所以具有较高的土壤含水量, 最大值可以达到79.19%, 进入6月份干旱季节土壤含水量逐渐下降, 最小值达到37.81%, 7月中旬以后进入雨季, 土壤含水量迅速升高并于8月23日达到最大值82.19%, 进入秋季之后缓慢降低并逐渐趋于平稳.在整个观测期间5个凋落物处理下土壤含水量平均值分别为62.93%(NL+NH)、63.28%(NL)、63.13%(CK)、63.22%(DL)和62.86%(DL+DH).不同凋落物处理并没有对小叶章湿地土壤含水量造成显著影响, 这可能是受到沼泽湿地的季节性积水影响.
 |
| 图 4 小叶章湿地土壤含水量季节动态 Fig. 4 Seasonal variations of soil watercontent in Calamagrostis angustifolia wetland |
5个凋落物处理小叶章湿地CO2排放通量均表现明显的季节性变化特点(图 5a), CO2排放通量均是在春季较低, 随着时间变化到夏季出现最高值, 之后随着时间逐渐降低并趋于平稳.不同处理间CO2排放通量季节变化范围分别为24.51~750.53(NL+NH)、27.92~771.72(NL)、57.53~816.74(CK)、62.31~867.82(DL)和66.33~911.72 mg·m-2·h-1(DL+DH), 平均排放通量分别为308.53、333.22、367.91、407.73和445.92 mg·m-2·h-1, 具体表现为(DL+DH)处理>(DL)处理>(CK)处理>(NL)处理>(NL+NH)处理.与自然对照样地相比, 添加凋落物显著地增加了小叶章湿地生长季CO2排放通量, 去除凋落物显著地降低了小叶章湿地生长季CO2排放通量(p < 0.05, 图 6a).
 |
| 图 5 不同凋落物处理下小叶章湿地CO2、CH4、N2O排放通量时间动态(a, b, c分别表示CO2、CH4、N2O排放通量) Fig. 5 CO2、CH4、N2O temporal emissions flux of Calamagrostis angustifolia wetland under different litter treatment |
 |
| 图 6 不同凋落物处理下小叶章湿地CO2、CH4、N2O排放通量(a, b, c分别表示CO2、CH4、N2O排放通量) Fig. 6 CO2, CH4, N2O emissions flux of Calamagrostis angustifolia wetland underdifferent litter treatment |
5个凋落物处理小叶章湿地CH4排放通量均表现明显的季节性变化特点(图 5b).CH4排放通量在8月之前一直趋于稳定, 8月之后进入雨季开始迅速上升, (NL+NH)处理在8月23日达到最大值51.92 mg·m-2·h-1, 进入9月份迅速下降并逐渐趋于稳定.不同处理间CH4排放通量季节变化范围分别为-0.0467~51.92(NL+NH)、-0.0448~50.41(NL)、-0.9483~33.52(CK)、-0.0127~39.14(DL)和-0.3469~37.82 mg·m-2·h-1(DL+DH), 平均排放通量分别为10.12、8.91、6.13、6.71和7.42 mg·m-2·h-1, 具体表现为(NL+NH)处理>(NL)处理>(DL+DH)处理>(DL)处理>(CK)处理.与自然对照样地相比, 添加凋落物和去除凋落物均增加了小叶章湿地生长季CH4排放通量, 并且生长季CH4排放通量对去除凋落物的响应更加剧烈(p < 0.05, 图 6b).
5个凋落物处理小叶章湿地N2O排放通量均表现明显的季节性变化特点(图 5c).均是在春季出现较弱的吸收现象, 随着时间变化逐渐由吸收转变成排放, 并且在7月份和8月份分别达到排放最大值, 进入9月份逐渐下降并趋于稳定.不同处理间N2O排放通量季节变化范围分别为-0.0435~0.0879(NL+NH)、-0.0229~0.0982(NL)、-0.0428~0.1352(CK)、-0.0403~0.1484(DL)和-0.0131~0.1214 mg·m-2·h-1(DL+DH), 平均排放通量分别为0.0269、0.0261、0.0314、0.0354和0.0407 mg·m-2·h-1, 具体表现为(DL+DH)处理>(DL)处理>(CK)处理>(NL+NH)处理>(NL)处理.与自然对照样地相比, 添加凋落物显著的增加了小叶章湿地生长季N2O排放通量, 剔除凋落物显著的降低了小叶章湿地生长季N2O排放通量(p < 0.05, 图 6c).
3.4 生长季温室气体排放通量与土壤温度和含水量之间的关系通过图 7可以看出, 5个凋落物处理下土壤温度与CO2排放通量之间均呈显著指数相关关系, 回归方程系数如表 2所示.土壤温度在5个凋落物处理中分别解释了72.41%、74.13%、75.42%、71.35%和75.27% CO2排放通量的变异.与此同时, 5个凋落物处理下土壤温度与CH4和N2O排放通量之间相关性不显著(图 7), 说明土壤温度不能很好地解释凋落物处理下CH4和N2O排放通量.5个凋落物处理下土壤含水量与CO2、CH4和N2O排放通量之间没有出现显著相关(图 7).这可能由于小叶章湿地季节性积水导致土壤含水量不能很好的解释小叶章湿地温室气体排放通量.
 |
| 图 7 不同凋落物处理下小叶章湿地CO2、CH4、N2O排放通量与土壤温度和含水量的回归关系(注:a, b, c分别表示CO2、CH4、N2O排放通量与土壤温度回归关系; d, e, f分别表示CO2、CH4、N2O排放通量与土壤含水量回归关系.) Fig. 7 Regressionrelationshipsbetween CO2, CH4, N2Oemissions flux, soil temperature and moistureof Calamagrostis angustifolia wetland underdifferent litter treatment |
| 表 2 不同凋落物处理CO2排放通量与土壤温度的指数关系模型 Table 2 Exponential relationships between CO2 flux and soil temperature under different litter treatments |
生态系统呼吸作用决定了CO2排放过程, 呼吸作用主要包括自养呼吸和异养呼吸两部分(窦景鑫等, 2009;李娇等, 2014).生态系统呼吸主要受非生物因子(温度、水分、土壤养分等)和生物因子(土壤动物和土壤微生物)两方面的影响.本研究中5个凋落物处理下的CO2排放均主要集中在7、8月份, 这主要是因为这个时期小叶章湿地大气温度、土壤温度以及降雨量均是整个生长季的高峰期, 提高了土壤微生物活性, 从而增强了呼吸作用, 使得CO2排放增加, 这与前人的研究结论基本一致(宋长春等, 2006;张荣涛等, 2018).凋落物是土壤有机质的重要来源, 不同凋落物处理可以改变土壤微环境、土壤养分以及凋落物初始质量, 进一步对土壤微生物群落结构和土壤呼吸产生影响(Crow et al., 2009; Chen et al., 2012).目前对凋落物去除和添加对生态系统影响的研究主要集中在土壤微生物结构和功能、土壤呼吸、土壤氮矿化等方面, 但主要在森林和草原生态系统中开展相关研究.张灏等(2014)通过对亚热带森林地表CO2通量对凋落物添加和去除响应的研究发现, 去除凋落物显著降低了森林CO2通量, 而添加凋落物增加了森林CO2通量;高强等(2016)通过对木荷林土壤呼吸对凋落物添加和去除响应的研究发现, 凋落物去除显著的降低了土壤呼吸, 但凋落物添加处理却没有对土壤呼吸造成显著影响;张素彦等(2018)通过对草原生态系统的研究发现, 添加凋落物显著的增加了草原生态系统碳通量.而凋落物去除和添加对湿地生态系统温室气体通量的影响报道很少.本研究结果显示, 凋落物添加显著的促进了小叶章湿地CO2排放, 去除凋落物也显著的降低了湿地CO2排放, 与前人研究结果类似.这可能是由于凋落物添加增加了微生物可利用的活性碳源, 使得微生物可以更好的分解利用有机碳.另外, 新鲜凋落物的输入刺激土壤中原有有机质的分解, 即“激发效应”(Kuzyakov et al., 2010;王清奎, 2011), 也会影响土壤呼吸.去除凋落物在减少地表碳源的同时也改变了土壤水热条件, 从而影响了土壤呼吸(Leff et al., 2012; Wang et al., 2013;熊莉等, 2015).本研究中, 不同凋落物处理下CO2排放通量与土壤10 cm温度出现了显著指数相关关系, 这可能是由于土壤温度影响了土壤微生物活性从而进一步改变了CO2排放, 因此本研究中凋落物添加和去除对CO2排放的影响主要是由于凋落物添加和去除处理改变了土壤微生物活性, 与前人的研究成果基本一致(Leroy et al., 2017; Xie et al., 2017).
CH4的排放通量主要由CH4产生和氧化过程之间的相互作用所决定(牟晓杰, 2012).凋落物是土壤碳的主要来源, 因此, 凋落物不仅对土壤CO2产生影响, 也可能会对土壤CH4有较大影响.凋落物层也可通过改变小气候和土壤理化性质来影响碳循环, 从而对土壤CH4排放产生重要的影响.本研究结果发现凋落物添加显著的增加了小叶章湿地CH4排放通量, 这与前人研究结果基本一致(Xie et al., 2010; Kayranli et al., 2010; Yavitt et al., 2015).这主要是由于凋落物添加后, 土壤中微生物可利用基质增加;同时增加的还有土壤中有机质以及可溶性有机碳含量(Strack et al., 2007张雪雯等, 2014).有机质含量变化也会影响土壤的氧化还原电位.相关研究表明, 添加凋落物后使得土壤氧化还原电位降低, 从而增加了CH4排放通量(Supparattanapan et al., 2010; Gaihre et al., 2013).另外, 本研究中去除凋落物也增加了小叶章湿地CH4排放通量, 这可能主要是因为湿地季节性积水的原因, 在土壤含水量较高的情况下(大于15.8%, 体积分数), 凋落物层增加了土壤对大气CH4的吸收;但在土壤含水量低的条件下, 凋落物层仅作为物理屏障, 这可能是因为凋落物层阻止了土壤和大气间的气体交换, 这与前人的研究结果一致(Price et al., 2003; Peichl et al., 2010; Wang et al., 2013).本研究结果发现, 5种凋落物处理下的小叶章湿地CH4排放均随着时间发生显著变化, 均是在夏季7、8月出现了排放的高峰期, 这主要包括两方面原因, 一方面由于植物在夏季生长旺盛, 自身功能以及植物通气组织数量都处在最佳状态, 另一方面因为研究区域存在季节性积水, 使得7、8月份积水期CH4排放远高于其它未积水时期.这种季节性积水带来的影响也表明, 无论是添加或是去除凋落物处理, 水分条件仍然是小叶章湿地CH4排放的主要影响因子.
生态系统N2O排放主要由土壤硝化和反硝化作用决定(王改玲等, 2010).凋落物不仅是土壤碳的主要来源, 同时也是土壤氮的主要来源, 不同凋落物处理对N2O排放过程的影响主要是源于凋落物输入后改变了土壤C/N以及影响了湿地土壤硝化和反硝化进程(Stapleton et al., 2005; Liu et al., 2008).另外, 凋落物层本身也可通过改变土壤水热因子来影响氮循环, 从而对土壤N2O排放产生重要的影响.Wang等(2014)在亚热带松树林发现去除凋落物土壤N2O排放通量降低了15%;Marhan等(2015)通过室内实验发现添加凋落物显著的增加了N2O排放通量.在本研究中, 小叶章湿地N2O排放通量随着添加凋落物显著增加;去除凋落物也显著的降低了N2O排放通量, 这与前人结果一致.这可能是由于, 凋落物会通过供应N、气体扩散和N2O的消耗来影响土壤-大气间的N2O通量.首先, 地上凋落物的矿化会提供充足的矿物N, 因此可能会促进土壤N2O的产生(Brüggemann et al., 2005; Zhang et al., 2008; Eickenscheidt et al., 2013).那么去除凋落物就可能会降低土壤-大气间的N2O交换.而且, 凋落物层作为物理屏障也会阻止大气和土壤之间的气体(如O2和N2O)扩散.但也有研究表明, 亚热带和热带森林凋落物去除对土壤-大气间N2O通量没有显著影响, 这表明N2O的产生主要和矿质土壤相联系, 而不是和土壤表面的凋落物层有关(Tang et al., 2006; Yan et al., 2008).这也体现出在不同的生态系统中, 凋落物对土壤N2O排放影响的作用机制不尽相同.本研究中N2O排放通量未与土壤温度和土壤含水量出现显著性相关, 这可能由于温室气体排放通量的影响因素较为复杂, 而且湿地生态系统中N2O排放过程中起主导作用的微生物群落和功能是如何响应凋落物处理及其内在驱动机制, 也将成为作者下一步研究的重点.
5 结论(Conclusions)1)(DL+DH、DL)处理增加了CO2、CH4和N2O排放通量;(NL+NH、NL)处理降低了CO2、N2O排放通量的同时增加了CH4排放通量.
2) 添加和去除凋落物均未改变CO2、CH4和N2O排放的季节动态.CO2排放通量与土壤温度出现了极显著正相关关系, CH4和N2O排放通量未与土壤温度出现显著相关关系.CO2、CH4和N2O排放通量均未与土壤含水量出现显著相关关系.
3) 未来还需进行长期试验验证, 揭示湿地凋落物对温室气体排放调节的内在机制, 既可以为湿地生态系统碳氮循环过程的研究提供理论基础, 也可以为未来气候变化中湿地生态系统的科学管理和生态保护提供有价值的参考.
Brüggemann N, Rosenkranz P, Papen H, et al. 2005. Pure stands oftemperate forest tree species modify soil respiration and N turnover[J]. Biogeoscience Discuss, 2: 303-331. DOI:10.5194/bgd-2-303-2005 |
Crow S E, Lajtha K, Filley T R, et al. 2009. Sources of plant-derived carbon and stability of organic matter in soil:Implications for global change[J]. Global Change Biology, 15: 2003-2019. DOI:10.1111/j.1365-2486.2009.01850.x |
Chen D, Zhou L, Wu J, et al. 2012. Tree girdling affects the soil microbial community by modifying resource availability in two subtropical plantations[J]. Applied Soil Ecology, 53: 0-115. |
窦晶鑫, 刘景双, 王洋, 等. 2009. 三江平原草甸湿地土壤有机碳矿化对C/N的响应[J]. 地理科学, 29(5): 773-778. |
Eickenscheidt N, Brumme R. 2013. Regulation of N2O and NOx emission patterns in six acidtemperate beech forest soils by soil gas diffusivity, N turnover, and atmospheric NOx concentrations[J]. Plant Soil, 369: 515-529. DOI:10.1007/s11104-013-1602-7 |
Gaihre Y K, Wassmann R, Villegas-Pangga G. 2013. Impact of elevated temperatures on greenhouse gas emissions in rice systems:interaction with straw incorporation studied in a growth chamber experiment[J]. Plant and Soil, 373: 857-875. DOI:10.1007/s11104-013-1852-4 |
高强, 马明睿, 韩华, 等. 2016. 去除和添加凋落物对木荷林土壤呼吸的短期影响[J]. 生态学杂志, 34(5): 1189-1197. |
IPCC. 2007. Climate change 2007: The physical science basis: Contribution of Working Group I tothe Fourth Assesment Report of the Intergovernmental Panel on Climate Change. CambridgeUniversity Press, Cambridge, United Kingdom and New York: NY, USA 996
|
Kayranli B, Scholz M, Mustafa A, et al. 2010. Carbon storage and fluxes within freshwater wetlands:A critical review[J]. Wetlands, 30: 111-124. DOI:10.1007/s13157-009-0003-4 |
Kuzyakov Y. 2010. Priming effects:Interactions between living and dead organic matter[J]. Soil Biology & Biochemistry, 42: 1363-1371. |
Liu D Y, Song C C, Wang L, et al. 2008. Exogenous nitrogen enrichment impact on the carbon mineralization and DOC of the freshwater marsh soil[J]. Environmental Science, 29: 3525-3530. |
Leff J W, Wieder W R, Taylor P G, et al. 2012. Experimental litterfall manipulation drives large and rapid changes in soil carbon cycling in a wet tropical forest[J]. Global Change Biology, 18: 2969-2979. DOI:10.1111/j.1365-2486.2012.02749.x |
Leroy F, Gogo S, Guimbaud C, et al. 2017. Vegetation composition controls temperature sensitivity of CO2 and CH4 emissions and DOC concentration in peatlands[J]. Soil Biology and Biochemistry, 107: 164-167. DOI:10.1016/j.soilbio.2017.01.005 |
Mearns L O, Katz R W, Schneider S H. 1984. Changes in the probabilities of extreme hightemperature events with changes in global mean temperature[J]. Journal of Climate and AppliedMeteorology, 23: 1601-1613. |
李娇, 尹春英, 周晓波, 等. 2014. 施氮对青藏高原东缘窄叶鲜卑花灌丛土壤呼吸的影响[J]. 生态学报, 34(19): 5558-5569. |
Marhan S, Auber J, Poll C. 2015. Additive effects of earthworms, nitrogen-rich litter and elevated soil temperature on N2O emission and nitrate leaching from an arable soil[J]. Applied Soil Ecology, 86: 55-61. DOI:10.1016/j.apsoil.2014.10.006 |
Niu S, Yang H, Zhang Z, et al. 2009. Non-additive effects of water and nitrogen addition on ecosystem carbon exchange in a temperate steppe[J]. Ecosystems, 12: 915-926. DOI:10.1007/s10021-009-9265-1 |
Price S J, Sherlock R R, Kelliher F M, et al. 2004. Pristine New Zealand forest soil is a strong methane sink[J]. Global Change Biology, 10(1): 16-26. DOI:10.1046/j.1529-8817.2003.00710x |
Peichl M, Arain M A, Ullah S, et al. 2010. Carbon dioxide, methane, and nitrous oxide exchanges in an age-sequence of temperate pine forests[J]. Global Change Biology, 16: 2198-2212. |
秦晓波, 李玉娥, 刘克樱, 等. 2006. 不同施肥处理稻田甲烷和氧化亚氮排放特征[J]. 农业工程学报, (7): 143-148. DOI:10.3321/j.issn:1002-6819.2006.07.030 |
Stapleton L M, Crout N M J, Säwström C, et al. 2005. Microbial carbon dynamics in nitrogen amended arctic tundra soil:Measurement and model testing[J]. Soil Biology & Biochemistry, 37: 2088-2098. |
Sayer E J, Tanner E V J, Lacey A L. 2006. Effects of litter manipulation on early-stage decomposition and meso-arthropod abundance in a tropical moist forest[J]. Forest Ecology and Management, 229: 285-293. DOI:10.1016/j.foreco.2006.04.007 |
宋长春, 张丽华, 王毅勇, 等. 2006. 淡水沼泽湿地CO2、CH4和N2O排放通量年际变化及其对氮输入的响应[J]. 环境科学, 27(12): 2369-2375. DOI:10.3321/j.issn:0250-3301.2006.12.001 |
Strack M, Waddington J M. 2007. Response of peatland carbon dioxide and methane fluxes to a water table drawdown experiment[J]. Global Biogeochemical Cycles, 21: 1-13. |
Supparattanapan S, Sejan P, Quantin C, et al. 2010. Salinity and organic amendment effects on methane production of a rain-fed saline paddy field[J]. Soil Science & Plant Nutrition, 55: 142-149. |
Sayer E J, Heard M S, Grant H K, et al. 2011. Soil carbon release enhanced by increased tropical forest litterfall[J]. Nature Climate Change, 1: 304-307. DOI:10.1038/nclimate1190 |
Sangsub C, Hee-Myung C, Sang-Hoon L, et al. 2017. Effect of elevated atmospheric CO2 concentration on growth and leaf litter decomposition of Quercus acutissima and Fraxinus rhynchophylla[J]. Plos One, 12: e0171197. DOI:10.1371/journal.pone.0171197 |
Tang X L, Liu S G, Zhou G Y, et al. 2006. Soil-atmospheric exchange of CO2, CH4, and N2O in three subtropical forest ecosystems in southern China[J]. Global Change Biology, 12: 546-560. DOI:10.1111/j.1365-2486.2006.01109.x |
Wickland K P, Striegl R G, Mast M A, et al. 2001. Carbon gas exchange at a southern Rocky Mountain wetland, 1996-1998[J]. Global Biogeochemical Cycles, 15: 321-335. DOI:10.1029/2000GB001325 |
Wang J B, Zhu T C, Ni H W, et al. 2013. Effects of elevated CO2 and nitrogen deposition on ecosystem carbon fluxes on the Sanjiang plain wetland in Northeast China[J]. PLOS ONE, 8: e66563. DOI:10.1371/journal.pone.0066563 |
Wang Y D, Wang H M, Ma Z Q, et al. 2013. The litter layer acts as a moisture-induced bidirectional buffer for atmospheric methane uptake by soil of a subtropical pine plantation[J]. Soil Biology and Biochemistry, 66: 45-50. DOI:10.1016/j.soilbio.2013.06.018 |
Wang Y D, Wang H M, Wang Z L, et al. 2014. Effect of litter layeron soil-atmosphere N2O flux of a subtropical pine plantation in China[J]. Atmospheric Environment, 82: 106-112. DOI:10.1016/j.atmosenv.2013.10.028 |
王改玲, 陈德立, 李勇. 2010. 土壤温度、水分和NH4+-N浓度对土壤硝化反应速度及N2O排放的影响[J]. 中国生态农业学报, 18(1): 1-6. |
王清奎. 2011. 碳输入方式对森林土壤碳库和碳循环的影响研究进展[J]. 应用生态学报, 22(4): 1075-1081. |
Xie Z B, Liu G, Bei Q C, et al. 2010. CO2mitigation potential in farmland of China by altering current organic matter amendment pattern[J]. Earth Science, 53: 1351-1357. |
熊莉, 徐振锋, 杨万勤, 等. 2015. 川西亚高山粗枝云杉人工林地上凋落物对土壤呼吸的贡献[J]. 生态学报, 35(14): 4678-4686. |
Xie H H, Ma W Y, Zhao C Y, et al. 2017. Effects of litter and moss on soil respiration in a spruce forest in the Qilian Mountains, Qinghai[J]. Acta Ecologica Sinica, 37(5): 1379-1390. |
Yan Y P, Sha L Q, Cao M, et al. 2008. Fluxes of CH4 and N2O from soil under a tropical seasonal rain forest inXishuangbanna, Southwest China[J]. Journal of Environmental Sciences, 20: 207-215. DOI:10.1016/S1001-0742(08)60033-9 |
Yavitt J B, Williams C J. 2015. Conifer litter identity regulates anaerobic microbial activity in wetland soils via variation in leaf litter chemical composition[J]. Geoderma, 243-244: 141-148. DOI:10.1016/j.geoderma.2014.12.023 |
Zhang W, Mo J M, Yu G R, et al. 2008. Emissions of nitrous oxidefrom three tropical forests in Southern China in response to simulated nitrogen deposition[J]. Plant and Soil, 306: 221-236. DOI:10.1007/s11104-008-9575-7 |
张雪雯, 莫熠, 张博雅, 等. 2014. 干湿交替及凋落物对若尔盖泥炭土可溶性有机碳的影响[J]. 湿地科学, (2): 134-140. |
张艺, 王春梅, 许可, 等. 2016. 若尔盖湿地土壤温室气体排放对模拟氮沉降增加的初期响应[J]. 北京林业大学学报, 38(8): 54-63. |
张素彦, 蒋红志, 王扬, 等. 2018. 凋落物去除和添加处理对典型草原生态系统碳通量的影响[J]. 植物生态学报, 42(3): 95-106. |
张荣涛, 隋心, 许楠, 等. 2018. 三江平原小叶章湿地温室气体排放及其对模拟氮沉降的响应[J]. 应用生态学报, 29(10): 35-42. |