 2020, Vol. 40
2020, Vol. 40


2. 二氧化碳捕集与封存国家与地方工程研究中心, 西安 710000
2. National & Local Joint Engineering Research Center of Carbon Capture and Storage Technology, Xi'an 710000
在全球大气环境CO2浓度不断升高的背景下, 发展新兴、高效、大规模的碳减排技术迫在眉睫.碳捕集与封存(CO2 capture and storage, CCS)技术被认为是目前最具潜力和优势的大规模排放源CO2捕集与长期储存的技术(Romanov et al., 2009;Marie et al., 2014), CCS技术能够将CO2从工业生产集中排放源中分离出来, 通过压缩、运输, 最终注入到深部咸水层、开采中或已废弃的油气层等稳定的地质构造中, 以实现CO2与大气的长期隔绝(Narita et al., 2012), 为以化石燃料和碳排放密集型产业在经济发展中持续发挥作用, 提供了有效的碳减排手段(纪翔等, 2018), 为减少CO2排放、缓减全球气候变化, 实现可持续发展提供了有力的技术支撑(Anwar et al., 2018;陈新新等, 2019).但在实际CCS项目操作中, 由于技术和地质条件变化等不稳定因素的影响, 仍存在一定的泄漏风险, 封存的CO2可通过跨越水力圈闭或盖层孔隙等途径发生慢速泄漏, 也可通过裂隙、断层、废弃井、未封闭处理的井眼或沿着井套发生快速泄漏(中国二氧化碳地质封存环境风险研究组, 2018), 从而对泄漏区生态系统产生严重影响.因此, 开展地质封存CO2泄漏对生态环境的影响研究就具有十分重要的实践意义.
目前有关CCS项目CO2泄漏对生态环境的影响较为广泛, 其中对土壤组成、理化性质(伍洋等, 2012;Wei et al., 2012;聂莉娟等, 2015)、微生物群落(Oppermann et al., 2010;陈浮等, 2017;张旺园等, 2017)、土壤酶活性(邓红章等, 2017)等土壤环境影响研究以及CO2泄漏对玉米、小麦、高粱、黑麦草、豌豆、大豆等旱作作物的生长形态、光合作用、植物酶的影响研究较多(Pierce et al., 2009;Ravi et al., 2010;Eline et al., 2011;蒋金豹等, 2013), 而对水环境的影响尤其是对地表水环境的影响鲜有报道.
水稻作为我国主要的粮食作物, 种植面积达340多万hm2, 占全国耕地的27%(李奕霏等, 2018), 其主产区域集中在东北地区、长江流域和珠江流域的平原区和盆地区.而广泛分布于沉积盆地中的深部咸水层, 是无大规模煤层和油气田地区主要的CO2地下储存场地(郑菲等, 2014).我国四川盆地大部、华北平原大部、苏北盆地以及准格尔盆地东南部, 因其深部咸水层储量大, CO2排放源密集、排放量大, 而成为将来优先考虑的CO2封存场地(李小春等, 2006).并且水稻作为水生作物, 其田间管理方式与旱作区别显著, 因此研究CO2泄漏对水稻的生存环境及其生长发育的影响具有重要意义.以水稻箱体种植为对象, 以一定速率CO2气体通入为控制, 是模拟地质封存CO2泄漏对水稻、稻田水及水稻土影响研究的有效方法.本研究以水稻箱体室外种植实验为对象, 探讨不同CO2泄漏速率对稻田水基础水质的影响规律, 为通过稻田水水质变化开展CO2泄漏监测提供依据.
2 材料与方法(Materials and methods) 2.1 实验作物、土壤与灌溉水本实验场地位于西北大学长安校区室外实验用地的塑料雨棚内.实验水稻选用当地常见的龙粳31号和龙稻18号两个品种, 其生育期为130~135 d, ≥10 ℃积温应大于2350 ℃.实验土壤来自学校周边的闲置农田土壤, 其理化性质见表 1.灌溉用水为自来水曝晒3 d后的花达水, 水质指标见表 2.
| 表 1 实验土壤基本理化性质 Table 1 Physical and chemical properties of experimental soil |
| 表 2 实验灌溉水水质指标 Table 2 Water quality indexes of experimental irrigation water |
本实验装置包括:CO2气罐、减压阀、流量计、分流器、曝气头、通气管及种植箱(图 1), 其中种植箱为70 cm×48 cm×45 cm的透明塑料箱.
 |
| 图 1 实验装置图 Fig. 1 The experimental device |
在减压阀和流量计控制下, 将气罐中的CO2气体由通气总管经分流器分流到4个支气管至曝气头, 通入箱体底部砾石层中, 曝气头固定于箱体底部长宽四等分点处.箱体底部敷设5 cm厚的粒径为5~10 mm花岗岩砾石层作为布气层;砾石层上覆20目乙烯网, 以保障布气层的通气性;在乙烯网上布设30 cm土层作为水稻生长的基质;种植中保障土层上覆水层厚度为4~6 cm.
实验种植龙粳31号和龙稻18号两个水稻品种, 设3个平行组, 共计30个种植箱, 每个品种包括1个对照组(CK)和4个CO2泄漏速率分别为0.06、0.12、0.24及0.48 L·m-2·min-1的实验组.
实验用水稻秧苗于2019年5月6日下种, 5~7 d出苗, 育苗时间共30 d.实验于6月5日施用复合肥(w(N):w(P2O5):w(K2O)=15%:15%:15%)作为基肥, 施肥标准为37.5 g·m-2, 然后用花达水进行灌溉.6月6日插秧, 插秧规格为30 cm×12.5 cm, 共种植9穴, 每穴4株.7月6日开始24 h持续通气, 直至9月28日停气、收获, 总通气时间84 d.其它如水分管理、病虫害防治等管理措施与大田一致.
2.3 指标测定与数据处理实验采用FC-100多功能水质检测仪(华东理工大学, 上海), 以对照组和实验组箱体长宽四等分点处的4个稻田水水样的混合样为样品, 现场测定其溶解性CO2(DCO2)、pH、溶解氧(DO)和氧化还原电位(ORP)等4项水质指标, 以反映不同CO2泄漏速率下CO2对稻田水水质的影响.
数据处理、统计分析与图表制作采用Microsoft Office Excel 2010、SPSS 22.0和origin9.5软件.
3 结果与分析(Results and analysis) 3.1 CO2泄漏对稻田水质的长期影响稻田水和稻田土壤作为水稻生长发育的基础条件, 其水质特征直接关系着水稻所需营养物质在土壤中的状态及其有效性, 也关系着水稻根系及地上部分的生长发育.CO2作为弱酸性气体, 当其泄漏进入水体时, 会导致部分CO2溶解在水中, 直接或间接影响稻田水的溶解性CO2(DCO2)、pH、溶解氧(DO)及氧化还原电位(ORP)等水质指标的变化.
3.1.1 CO2泄漏对稻田水溶解性CO2(DCO2)的影响当CO2溶于水中后, 又会因CO2-H2CO3-HCO3--CO32-体系平衡的移动而导致其存在状态的变化.常把CO2和H2CO3合并为H2CO3*来表示水中CO2含量, 因水中H2CO3含量极低, 该指标主要指溶解性气体CO2(戴树桂, 2006), 本文称其为溶解性CO2(DCO2).
在不同CO2泄漏速率下, 龙粳31和龙稻18两个品种水稻的稻田水DCO2随时间的变化如图 2所示.由图可知, 在一定CO2速率下, DCO2随时间先迅速上升, 一定时间后缓慢升高并趋于平缓, 最终达到相对稳定状态;随着CO2泄漏速率的增加, DCO2值升高幅度越来越大.通过模拟不同CO2泄漏速率下DCO2随时间的变化规律发现, 指数函数能够很好地拟合DCO2随时间的变化, 不同CO2速率下DCO2随时间变化的拟合方程的R2为0.9768~0.9977(n=11), 均达到了极显著水平.由拟合方程可知, DCO2随时间变化的最终平衡值为拟合方程的常数项.以拟合方程的最终平衡值为基础, 以拟合方程的拟合值与最终平衡值相对误差的1%为标准, 确定DCO2平衡时间, 求取不同CO2速率下的DCO2平衡时间(表 3).由表可知, 当CO2泄漏速率从0.06 L·m-2·min-1增加到0.48 L·m-2·min-1, 龙粳31稻田水的DCO2平衡值由272.39 mg·L-1上升到619.64 mg·L-1, 达到平衡的时间从67.47 h减少到39.14 h;龙稻18稻田水的DCO2平衡值从270.21 mg·L-1上升到594.92 mg·L-1, 达到平衡的时间从63.28 h减少到35.45 h.即DCO2的平衡值随着CO2泄漏速率的增加而增加, 达到平衡所需的时间随着CO2泄漏速率的增加而减少.
 |
| 图 2 不同CO2泄漏速率下稻田水DCO2随时间的变化(a.龙粳31, b.龙稻18) Fig. 2 The change of DCO2 in paddy field with time under different CO2 leakage rates |
| 表 3 稻田水DCO2在不同CO2泄漏速率下的拟合平衡值与平衡时间 Table 3 Fitting equilibrium value and equilibrium time of DCO2 in paddy field with different CO2 leakage rates |
根据DCO2拟合平衡时间, 利用SPSS软件对CK组及各处理组基本稳定后的DCO2进行差异性分析(图 3).不同CO2处理组的龙粳31稻田水和龙稻18稻田水DCO2均与CK组之间存在极显著差异;与CK组相比, 不同速率CO2通入时, 龙粳31稻田水的DCO2分别上升了16.09倍、24.64倍、30.20倍、37.69倍, 龙稻18稻田水DCO2分别上升了22.25倍、30.99倍、42.56倍、50.22倍.
 |
| 图 3 不同CO2泄漏速率下稻田水质平衡后DCO2差异分析(p < 0.05)(a.龙粳31, b.龙稻18) Fig. 3 The difference of DCO2 after water quality balance in rice field with different CO2 leakage rates (p < 0.05) |
pH是水质的基础指标, 稻田水的pH能够控制和影响稻田土壤的矿物形态、水土界面交换、水体中多种体系的平衡, 影响土壤营养元素利用效率、土壤微生物的群落特征及水-土壤-根系的界面交换机制与过程(韦梦等, 2015;陈平平等, 2017).在不同CO2泄漏速率下, 两个品种水稻的稻田水pH随时间的变化如图 4所示.在一定CO2泄漏速率下, 稻田水pH值随时间先迅速下降, 一定时间后缓慢下降并趋于平缓, 最终达到相对稳定状态;随着CO2速率的增加, pH值降低幅度越来越大.采用与DCO2相同的方法对pH值进行拟合计算, 求取不同CO2速率下的pH平衡时间(表 4).当CO2泄漏速率从0.06 L·m-2·min-1增加到0.48 L·m-2·min-1时, 龙粳31稻田水的pH平衡值从6.58下降到6.38, 达到平衡的时间从36.89 h减少到9.72 h;龙稻18稻田水的pH平衡值从6.61下降到6.34, 达到平衡的时间从35.23 h减少到9.53 h.即pH的平衡值和达到平衡所需的时间随着CO2泄漏速率的增加而降低.
 |
| 图 4 不同CO2泄漏速率下稻田水pH随时间的变化(a.龙粳31, b.龙稻18) Fig. 4 The change of pH in paddy field with time under different CO2 leakage rates |
| 表 4 不同CO2泄漏速率下稻田水pH的平衡值与平衡时间 Table 4 Fitting equilibrium value and equilibrium time of pH in paddy field with different CO2 leakage rates |
根据pH拟合平衡时间, 对CK组及各处理组基本稳定后的pH进行差异性分析(图 5).不同速率CO2处理组的龙粳31稻田水pH均与CK组之间存在极显著差异;不同速率CO2处理组稻田水pH之间, 除0.24 L·m-2·min-1与0.48 L·m-2·min-1 CO2速率组之间差异不显著外, 其它组间差异显著.不同速率CO2处理组龙稻18稻田水pH均与CK组之间存在极显著差异;不同速率CO2处理组稻田水pH之间, 除0.24 L·m-2·min-1 CO2泄漏速率下的pH与0.12 L·m-2·min-1及0.48 L·m-2·min-1泄漏速率下的pH差异不显著外, 其余处理组之间差异显著.与CK组相比, 不同速率CO2通入时, 龙粳31稻田水的pH分别下降了20.22%、20.87%、22.32%、23.09%, 龙稻18稻田水的pH分别下降了20.44%、22.08%、23.16%、24.00%.
 |
| 图 5 不同CO2泄漏速率下稻田水质平衡后pH差异分析(p < 0.05)(a.龙粳31, b.龙稻18) Fig. 5 The difference of pH after water quality balance in rice field with different CO2 leakage rate (p < 0.05) |
水体溶解氧是衡量水体水质的重要指标, 在稻田氮素循环和水稻生长发育过程中发挥了重要作用, 土壤氧化还原电位、pH值、离子形态、微生物活动等均可能受到溶解氧的影响, 从而直接或间接影响稻田生态系统(胡继杰等, 2017).不同CO2泄漏速率下两个品种水稻稻田水DO随时间的变化如图 6所示, 可以看出, DO随CO2泄漏速率和时间的变化规律同pH的变化规律一致.根据拟合方程可得DO平衡时间(表 5), 当CO2泄漏速率从0.06 L·m-2·min-1增加到0.48 L·m-2·min-1, 龙粳31稻田水的DO平衡值从7.30 mg·L-1下降到4.29 mg·L-1, 达到平衡的时间从56.10 h减少到37.49 h;龙稻18的稻田水DO平衡值从7.14 mg·L-1下降到4.59 mg·L-1, 达到平衡的时间从60.90 h减少到42.72 h.即DO的平衡值和达到平衡所需的时间随着CO2泄漏速率的增加而降低.
 |
| 图 6 不同CO2泄漏速率下稻田水DO随时间的变化(a.龙粳31, b.龙稻18) Fig. 6 The change of DO in paddy field with time under different CO2 leakage rates |
| 表 5 不同CO2泄漏速率下稻田水DO的平衡值与平衡时间 Table 5 Fitting equilibrium value and equilibrium time of DO in paddy field with different CO2 leakage rates |
同样, 根据DO拟合平衡时间, 对稻田水质稳定后的DO值进行与pH和DCO2相同的分析(图 7), 两个品种水稻稻田水的DO值之间的差异显著性规律同DCO2一致, 两个品种水稻稻田水在各CO2泄漏速率下的DO值与CK组存在极显著差异, 其余各处理组间也存在显著性差异.各CO2速率处理下龙粳31稻田水DO值分别较CK组降低了11.90%、20.76%、34.55%、45.74%, 龙稻18稻田水DO值分别较CK组降低了12.71%、23.93%、33.55%、46.30%.
 |
| 图 7 不同CO2泄漏速率下稻田水质平衡后DO差异分析(p < 0.05)(a.龙粳31, b.龙稻18) Fig. 7 The difference of DO after water quality balance in rice field with different CO2 leakage rates (p < 0.05) |
不同CO2泄漏速率下两个品种水稻稻田水ORP随时间的变化如图 8所示, ORP随CO2泄漏速率和时间的变化规律同DCO2的变化规律一致.根据拟合方程可得ORP平衡时间(表 6), 当CO2速率从0.06 L·m-2·min-1增加到0.48 L·m-2·min-1时, 龙粳31的ORP平衡值从246.91 mV上升至267.08 mV达到平衡的时间从60.37 h减少到15.11 h;龙稻18的ORP平衡值从239.59 mV上升至260.24 mV, 达到平衡的时间从57.53 h减少到18.37 h.即ORP的平衡值随着CO2泄漏速率的增加而上升, 达到平衡的时间随CO2泄漏速率的增加而减少.
 |
| 图 8 稻田水ORP在不同CO2泄漏速率下的变化(a.龙粳31, b.龙稻18) Fig. 8 The change of ORP in paddy field with time under different CO2 leakage rates |
| 表 6 水稻水ORP在不同CO2泄露速率下的拟合平衡值与平衡时间 Table 6 Fitting equilibrium value and equilibrium time of ORP in paddy field with different CO2 leakage rates |
根据ORP差异性分析(图 9)可知, 两个品种水稻的ORP之间的差异显著性规律不尽相同.不同CO2速率下两个品种的稻田水的ORP均与CK组ORP稳定值存在极显著差异, 但龙粳31稻田水的ORP除0.24 L·m-2·min-1和0.48 L·m-2·min-1两个处理组之间的差异不显著外, 其余处理组之间差异显著;龙稻18稻田水的ORP除各相邻速率组之间的差异不显著外, 不相邻速率组之间差异显著, 该结果与龙稻18的稻田水pH差异性分析结果相近.与CK组相比, 通入不同速率CO2时, 龙粳31稻田水的ORP分别升高了40.30%、42.02%、47.72%、50.57%, 龙稻18稻田水的ORP分别升高了44.28%、49.70%、53.31%、58.93%.
 |
| 图 9 不同CO2泄漏速率下稻田水质平衡后ORP差异分析(p < 0.05)(a.龙粳31, b.龙稻18) Fig. 9 The difference of ORP after water quality balance in rice field with different CO2 leakage rates (p < 0.05) |
从长期数据来看, 在一定CO2通入速率条件下, 稻田水水质指标随时间的变化呈先急剧变化后稳定于特定值的指数变化规律.但短期观测发现, 除通气初期的急剧变化外, 在各水质指标稳定后, 还表现为一定的日变化规律.在所有指标全部平衡之后, 于7月10日8:00、12:00、16:00、20:00测定了两个品种稻田水的水质指标, 以龙粳31号品种稻田水为例, 各水质指标昼间变化如图 10所示.由图可知, 在不同的CO2通入速率条件下, 稻田水各水质指标均呈现明显的日变化规律, 其中DCO2和DO的日变化更为显著;DCO2呈早晚高、午间低的先减后增规律, 而pH、DO和ORP均呈早晚低、午间高的先增后减规律.
 |
| 图 10 不同CO2泄漏速率下龙粳31号稻田水水质指标的日变化特征(a. DCO2, b. pH, c. DO, d. ORP) Fig. 10 Diurnal variation of water quality indexes in paddy field of longjing 31 with different CO2 leakage rates |
根据对照组和不同速率CO2通入条件下两种水稻稻田水水质指标随时间的指数变化规律, 以对照组各指标均值和不同速率CO2通入条件下两种水稻稻田水各水质指标平衡值为样本, 综合计算稻田水各单项指标及其与CO2泄漏速率的相关系数(表 7).稻田水的DCO2、pH、DO、ORP之间以及与CO2泄漏速率之间均存在相关性, 其中DCO2、DO与CO2通入速率在p < 0.01水平下显著相关, ORP、pH与CO2通入速率在p < 0.05水平下显著相关;DCO2、ORP与CO2通入速率呈正相关关系, pH、DO与CO2通入速率呈负相关关系.
| 表 7 稻田水水质指标及其与CO2泄漏速率的相关系数 Table 7 Correlation coefficient among rice field water quality indexes and its with CO2 leakage rates |
进一步探讨各指标平衡值与CO2通入速率的关系(图 11), 发现稻田水各水质指标的平衡值与CO2通入速率均呈对数函数关系, 其中DCO2和ORP的平衡值随着CO2通入速率的增加呈上升趋势, 而pH和DO则呈下降趋势.
 |
| 图 11 CO2泄漏速率与稻田水各水质指标的关系 Fig. 11 Relationship between CO2 leakage rates and water quality indexes in paddy fields |
通过模拟CO2泄漏对稻田水水质指标的影响监测, 发现CO2泄漏对稻田水的相关水质指标影响显著.CO2作为模拟CCS泄漏通入稻田水的物质载体, 稻田水中的DCO2就与其通入速率具有显著的正相关关系.CO2作为可溶性气体, 其在水中的溶解度随温度的增高而降低(汤勇等, 2011), 受温度日变化的影响, 稻田水中DCO2浓度呈早晚高、午间低的先降后升的日变化规律.DCO2作为影响稻田水中其他水质指标的根源, 也直接和间接影响着其它水质指标的变化.
实验发现, 在旱作土壤改为水作土壤后, 与原有旱作土壤的7.56和灌溉水的7.63相比, 稻田水的pH为8.11~8.89, 其原因可能与在淹水还原条件下, 旱作土壤中原有硝态氮大量还原减少有关(Narteh et al. 1999;唐罗忠等, 2005).在通入CO2的条件下, 稻田水的pH随时间按照指数规律迅速降低, 并平衡于一定的水平;随着CO2通入速率的增加其平衡值越来越低, 平衡时间越来越短.其原因是随着CO2通入速率的增加, 促进了CO2-H2CO3-HCO3--CO32-平衡体系中由溶解性CO2向生成H2CO3的方向及由H2CO3向HCO3-解离的方向移动.随着CO2通入速率的增加和时间的延长, 稻田水由通气前的碱性水快速变化为中性水及弱酸性水, 这一结果与张丙华等(2016)的CO2对水体pH的影响研究结果一致.但与其对地表水的影响相比, 对稻田水pH的影响幅度较小, 平衡时间更长, 其主要原因与稻田水作为水-土共存体系, 土壤中的弱酸强碱盐、强酸弱碱盐、两性化合物及土壤胶体对pH的强大缓冲作用有关(赵烨等, 2011).稻田水中pH的早晚低、午间高的先升后降的日变化规律与DCO2的日变化规律密切相关.
就DO的长期变化而言, 在一定CO2通入速率条件下, 稻田水中的DO随时间的延长呈指数变化规律;随着CO2通入速率的增加, 稻田水中DO的平衡浓度逐渐降低, 平衡时间不断缩短.一方面, 通入CO2对水中原有DO具有一定的驱替作用(张丙华等, 2016), 这种驱替作用随着CO2通入速率的增加而增加, 使得稻田水中DO的平衡浓度也随之降低;另一方面, 作为水生作物, 为了适应长期淹水条件下根际的氧化条件和对土壤中养分的吸收, 水稻具有大量结构复杂的、完整的通气组织, 保证将叶片光合作用产生的氧气, 经氧气输导组织传送到水稻根系, 并通过根系组织, 将储存的氧气释放到根际, 满足其根系生理活动的需求(李奕林等, 2012;刘依依等, 2014).稻田水中DO的早晚低、午间高的先升后降的短期日变化规律与DCO2的日变化规律具有明显的负相关关系, 但与DCO2日变化相比, DO的日变化更为显著, 这一点与水稻的泌氧作用具有极为密切的关系(付融冰等, 2008).即在CO2通入条件下, 稻田水的DO长期变化主要受CO2通入速率的控制, 而其短期日变化主要体现在以CO2通入速率为控制基础的光合泌氧作用所控制.
长期CO2泄漏实验结果显示, 在通入CO2的条件下, 稻田水ORP随时间呈指数型递增直至平衡的变化规律;随着CO2通入速率的增加, 稻田ORP的平衡值逐渐增加, 平衡时间不断缩短.其原因为在开放性水体环境中, 水体的ORP与由水气界面交换所控制的DO及有H+参与的多种氧化还原体系控制的pH关系密切, 当水中的溶解氧与大气的氧气平衡时, 其关系可表示为(尹军等, 2006):
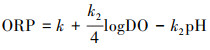
|
(1) |

|
(2) |
式中, R为气体常数, 8.314 J·mol-1·k-1;T为绝对温度(K);F为法拉第常数, 96485.309 C·mol-1;k为与温度有关的常数.
按照上述关系, 通过式(1)计算各种CO2泄漏速率下DO、pH及ORP等指标平衡值的k值(表 8), 由表可知, 各CO2泄漏速率下的k值为616.99~634.17, 其平均值为625.44, 各CO2泄漏速率下k值与k的平均值的相对误差仅为-1.40%~1.35%, k值接近于常数;再用k的平均值计算各实验组的ORP拟合值, 最后求得平衡值与拟合值的相对误差, 其相对误差变化于-3.38%~3.41%, 误差可以接受.因此, 在CO2泄漏条件下, 稻田水pH的大幅降低对稻田水ORP的增加作用显著大于DO减少对ORP的降低作用, 因此稻田水ORP整体呈上升趋势.对于稻田水的ORP早晚低、午间高的日变化规律, 可能与DO的日变规律关系更为密切, 这一结论与李华超等(2014)、陶敏等(2008)的研究结果一致.
| 表 8 不同CO2泄漏速率下稻田水ORP的平衡值、拟合值及相对误差 Table 8 Equilibrium value, fitting value and relative error of water ORP in paddy field with different CO2 leakage rate |
根据CO2泄漏条件下各指标与对照组之间差异性分析发现, 各泄漏条件下的龙粳31和龙稻18两个品种的稻田水DCO2、pH、DO、ORP均与CK组存在显著差异(表 9).对比发现, 4个基础水质指标中, 各种泄漏速率条件下DCO2为270.21 ~619.64 mg·L-1, 较实验CK组增长了16.09~50.22倍, 明显高于其它指标, 也明显高于Esteban等(2009)所报道的在CO2积累现象非常严重的循环水养殖系统的CO2浓度(42.9 mg·L-1).根据CCS类项目CO2风险泄漏的可能性, 建议将稻田水DCO2作为水稻种植区CO2泄漏风险监测的主要指标, 将pH、DO、ORP作为CO2泄漏监测的辅助指标.
| 表 9 稻田水水质指标较CK组变化幅度 Table 9 Amplitude of variation of water quality index in paddy field compared with CK |
1) 不同CO2泄漏速率下, 稻田水的DCO2、pH、DO和ORP等水质指标随时间的长期变化均呈指数函数关系, 其平衡时间随着CO2泄漏速率的增大而减小;CO2泄漏速率与稻田水水质指标相关性显著, 其中DCO2和ORP与CO2泄漏速率呈正相关, pH和DO与CO2泄漏速率呈负相关;CO2泄漏对稻田水质指标的影响与CK组差异显著, 不同CO2泄漏速率组之间DCO2和DO差异显著, pH和ORP差异较弱.
2) 在各指标平衡后, 不同CO2通入速率下, 稻田水各水质指标均呈现明显的日变化规律, 其中DCO2和DO的日变化更为显著;DCO2呈现早晚高、午间低的先减后增规律, 而pH、DO和ORP均呈现早晚低、午间高的先增后减规律.
3) 在CO2泄漏条件下各指标与对照组差异明显, 其中DCO2差异最为显著.建议将稻田水DCO2作为水稻种植区CO2泄漏风险监测的主要指标, 将pH、DO、ORP作为CO2泄漏监测的辅助指标.
Anwar M N, Fayyaz A, Sohail N F, et al. 2018. CO2 capture and storage:A way forward for sustainable environment[J]. Journal of Environmental Management, 226: 132-144. DOI:10.1016/j.jenvman.2018.08.009 |
陈浮, 杨宝丹, 马静, 等. 2017. 高浓度CO2地下泄漏对土壤微生物群落结构的影响[J]. 土壤学报, 54(1): 180-190. |
陈平平, 郭莉莉, 唐利忠, 等. 2017. 土壤pH对不同酸性敏感型水稻品种氮利用效率与根际土壤生物学特性的影响[J]. 核农学报, 31(4): 757-767. |
陈新新, 马俊杰, 李琦, 等. 2019. 国内地质封存CO2泄漏的生态影响研究[J]. 环境工程, 37(2): 27-34. |
戴树桂. 2006. 环境化学[M]. .
|
邓红章, 张慧慧, 李春荣, 等. 2017. 人工模拟地质封存CO2泄漏对土壤酶活性的影响[J]. 应用化工, 46(1): 4-9. |
Eline V, Dirk R, Patrick W. 2011. Considering sink strength to model crop production under elevated atmospheric CO2[J]. Agricultural and Forest Meteorology, 151(12): 1753-1762. |
Esteban J M E. 2009. Problems affecting nitrification in commercial RAS with fixed-bed biofilters for salmonids in Chile[J]. Aquacultural Engineering, 41(2): 91-96. |
付融冰, 朱宜平, 杨海真, 等. 2008. 连续流湿地中DO、ORP状况及与植物根系分布的关系[J]. 环境科学学报, 28(10): 2036-2041. |
胡继杰, 朱练峰, 钟楚, 等. 2017. 溶解氧对稻田土壤氮素转化及水稻氮代谢影响研究进展[J]. 生态学杂志, 36(7): 2019-2028. |
纪翔, 马欣, 韩耀杰, 等. 2018. 箱体模拟地质封存CO2泄漏速度差异对植物的影响[J]. 农业工程学报, 34(2): 242-247. |
蒋金豹, Steven M D, 何汝艳, 等. 2013. 利用大豆光谱特征判定地下封存CO2泄漏[J]. 农业工程学报, 29(12): 163-169. |
李华超, 陈宗晶, 陈章和. 2014. 六种湿地植物根际氧化还原电位的日变化[J]. 生态学报, 34(20): 5766-5773. |
李小春, 刘延锋, 白冰, 等. 2006. 中国深部咸水含水层CO2储存优先区域选择[J]. 岩石力学与工程学报, (5): 963-968. |
李奕霏, 肖谋良, 袁红朝, 等. 2018. CO2倍增对稻田土壤碳氮水解酶活性的影响[J]. 中国环境科学, 38(9): 3474-3480. |
李奕林. 2012. 水稻根系通气组织与根系泌氧及根际硝化作用的关系[J]. 生态学报, 32(7): 2066-2074. |
刘依依, 傅志强. 2014. 水稻根系泌氧特性及其影响因素[J]. 作物研究, 28(3): 312-315. |
Marie R. 2014. Carbon prices and CCS investment:A comparative study between the European Union and China[J]. Energy Policy, 75: 327-340. |
Narteh L T, Sahrawat K L. 1999. Influence of flooding on electrochemical and chemical properties of West African soils[J]. Geoderma, 87(3): 179-207. |
Narita D. 2012. Managing uncertainties:the making of the IPCC's special report on carbon dioxide capture and storage[J]. Public Understanding of Science, 21(1): 84-100. |
聂莉娟, 马俊杰, 赵雪峰, 等. 2015. 模拟CCS技术CO2泄漏对C3、C4作物土壤化学性质的影响[J]. 水土保持学报, 29(5): 200-205. |
Oppermann B I, Michaelis W, Blumenberg M, et al. 2010. Soil microbial community changes as a result of long-term exposure to a natural CO2 vent[J]. Geochemica Cosmochimica Acta, 74(9): 2697-2716. |
Pierce S, Sjogersten S. 2009. Effects of below ground CO2 emissions on plant and microbial communities[J]. Plant Soil, 325(1): 197-205. |
Ravi H P, Jeremy J C, Michael D S. 2010. Effects of CO2 gas as leaks from geological storage sites on agro-ecosystems[J]. Energy, 35(12): 4587-4591. |
Romanov V N, Ackman T E, Soong Y, et al. 2009. CO2 storage in shallow underground and surface coal mines:challenges and opportunities[J]. Environmental Science and Technology, 43(3): 561-564. DOI:10.1021/es801730v |
陶敏, 贺锋, 徐栋, 等. 2008. 复合垂直流人工湿地氧化还原特征及不同功能区净化作用研究[J]. 长江流域资源与环境, 17(2): 291-294. |
唐罗忠, 生原喜久雄, 户田浩人, 等. 2005. 湿地林土壤的Fe2+, Eh及pH值的变化[J]. 生态学报, (1): 103-107. |
汤勇, 杜志敏, 孙雷, 等. 2011. CO2在地层水中溶解对驱油过程的影响[J]. 石油学报, 32(2): 311-314. |
韦梦, 马兰, 胡越航, 等. 2015. 高温下两种不同pH水稻土甲烷产生及古菌群落的比较[J]. 生态学杂志, 34(6): 1667-1674. |
伍洋, 马欣, 李玉娥, 等. 2012. 地质封存CO2泄漏对农田生态系统的影响评估及耐受阈值[J]. 农业工程学报, 28(2): 196-205. |
Wei Y, Maroto-Valer M, Steven M D. 2011. Environment Consequences of Potential Leaks of CO2 in soil[J]. Energy Procedia, 4(2011): 3224-3230. |
尹军, 刘志生, 赵可, 等. 2006. 饮用水中无机成分与氧化还原电位的关系[J]. 环境与健康杂志, 23(2): 148-151. |
张丙华, 景炯炯, 耿春香, 等. 2016. 地质封存CO2泄露对地表水中非金属类指标的影响[J]. 水土保持通报, 36(2): 161-164+170. |
张旺园, 张绍良, 陈浮, 等. 2017. 模拟地下CO2泄漏对土壤微生物群落的短期影响[J]. 农业环境科学学报, 36(6): 1167-1176. |
赵烨, 等. 2011. 土壤环境科学与工程[M]. 北京: 北京师范大学出版社.
|
郑菲, 施小清, 吴吉春, 等. 2014. 深部咸水层CO2地质封存数值模拟参数的全局敏感性分析——以苏北盆地盐城组为例[J]. 吉林大学学报(地球科学版), 44(1): 310-318. |
中国二氧化碳地质封存环境风险研究组. 2018. 中国二氧化碳地质封存环境风险评估[M]. 北京: 化学工学出版社.
|