 2020, Vol. 40
2020, Vol. 40



溶解性有机质(dissolved organic matter, DOM)广泛存在于湖泊等水生态系统中, 其表生行为和环境效应十分丰富(吴丰昌, 2010; 何伟等, 2016).DOM是湖泊中非生物相和生物相之间的重要媒介, 并对污染物(如营养盐、重金属、有机污染物等)的生物地球化学循环和生态风险产生重要调控(雷沛等, 2018).湖泊沉积物是污染物主要的“汇”和“源”, 同时也是研究流域人类活动对湖泊水环境变化驱动的重要载体(Dubois et al., 2018).沉积物DOM是湖泊中普遍存在的活性组分, 其时空分布特征间接反映了湖泊污染物迁移转化规律, 也记录了湖泊及流域的演变信息(Ziegelgruber et al., 2013).
湖泊富营养化已成为世界范围内最严重的水环境问题之一, 蓝藻水华表现出爆发范围大、频率高、持续时间长的特点(Huisman et al., 2018).在一定条件下, 富营养化湖泊中的蓝藻在局部区域堆积, 厚度甚至达到数十厘米(尚丽霞等, 2013).高密度蓝藻堆积以后容易死亡并形成有机碎屑, 除小部分碎屑在水柱中被好氧异养微生物利用, 大部分有机碎屑会沉降到沉积物表面并发生分解, 导致沉积物中营养组分和微生物活性的增加.蓝藻会经历“复苏-爆发-衰亡-休眠”的生长循环, 这一过程会形成活性迥异的藻源性有机质并输送到沉积物中(孙共献, 2015), 是湖泊DOM最主要的来源(Zhang et al., 2009).例如, 蓝藻在生长期和衰亡期都会生成大量胞外聚合物(如多糖、蛋白质类)、难降解有机质(如腐殖质类)以及其它小分子酸类等物质(Villacorte et al., 2015), 这些物质会调控污染物的生物地球化学循环.关注不同藻华时期沉积物DOM性质, 对认识湖泊的富营养化控制和生态修复具有重要意义.
DOM是一类非均质混合物, 其组成和结构相对复杂(Chen et al., 2015).其中, 紫外-可见吸收光谱和三维荧光光谱具有灵敏度高、样品量少、样品预处理简单等特点(Pitta et al., 2017).近年来, 这两项技术已被广泛应用于DOM组分、来源研究中, 但大多数研究集中在湖库表层水、间隙水的光谱特征分析(Zhang et al., 2018; Shang et al., 2019; 陈乐等, 2019), 对受人为活动影响较大的富营养化湖泊主要关注沉积物固相有机质形态(赵萱等, 2012), 而针对不同藻华时期沉积物DOM性质鲜有报道.因此, 本研究选取典型富营养湖泊巢湖作为研究对象, 通过提取不同藻华时期沉积物DOM, 结合紫外-可见吸收光谱和三维荧光光谱解析, 揭示富营养化湖泊沉积物DOM时空分布规律, 为研究富营养化湖泊的环境演化和污染控制提供基础.
2 材料与方法(Materials and methods) 2.1 区域概况巢湖位于安徽中部, 为长江水系下游湖泊, 是我国第五大淡水湖.巢湖东西长54.5 km, 南北平均宽15.1 km, 最大水域面积约825 km2, 平均水深2.69 m.巢湖湖水补给主要来自地表径流, 沿湖共有河流35条, 呈放射状流入巢湖, 其中杭埠河、南淝河、白石天河3条河流入湖径流量占总径流量75%以上(Fang et al., 2017; 程红等, 2009).巢湖水体总体富营养化严重, 湖区内发生富营养化的水面面积约占全湖面积70%, 其中西半湖长期处于中度甚至重度富营养状态(姜霞等, 2010; 张民等, 2015), 呈现东低西高、湖周高、湖心低的特点(温胜芳2013).
2.2 样品采集与处理受东南季风的影响, 每年5—9月均有大量的蓝藻堆集于巢湖西半湖的北部湖湾(殷长秦等, 2012).因此, 选择该区域(L区)采集表层沉积物样品考察藻华堆集、沉降腐解对沉积物DOM的影响, 沿北湖滨带的湖周设置5个采样点, 分别为L1~L5;同时, 在西半湖的湖心区(C区)采集沉积物, 对比研究低藻密度堆积下沉积物DOM的空间差异, 选用网格布点法设置6个采样点, 分别为C1~C6, 各采样点位分布见图 1.在时间上, 对该研究区域一个藻华的循环周期, 包括复苏期(2018年4月, 春季)、爆发期(2018年8月, 夏季)、腐解期(2017年11月, 秋季)和休眠期(2019年1月, 冬季), 在相应点位分别开展表层沉积物采集, 以探讨沉积物DOM的季节差异.利用抓泥斗采集表层沉积物(0~10 cm), 同步采集湖泊表层水(水面0.5 m以下)500 mL以测定水体叶绿素a含量.此外, 在2018年8月夏季藻华爆发时, 利用自重力柱状采泥器(Corer 60, Uwitec, Austria)分别在C4和L5点采集柱状沉积物(柱长分别为30 cm和20 cm), 以探讨沉积物DOM的垂直分布特征.在现场利用分层装置将柱状样按2 cm进行分层.新鲜的表层沉积物样品以及分层的柱状样品置于4 ℃车载冰箱中尽快运回实验室.经20目筛网反复筛去砂石、贝类后冷冻干燥, 研磨并过100目筛后备用.
 |
| 图 1 巢湖沉积物采样点分布 Fig. 1 Sampling sites of western Chaohu Lake |
称取一定量的沉积物, 采用1 mol · L-1 KCl溶液按1 : 10 (质量体积比)提取沉积物中DOM (沈烁等, 2014; 翟天恩等, 2017; 高凤等, 2019).在恒温(20 ℃)、避光、振荡(200 r · min-1)条件下提取24 h.将提取后的样品进行离心(4000 r · min-1、15 min)获取上清液, 经0.45 μm滤膜过滤置于4 ℃待用.
2.3.2 水体、沉积物叶绿素a以及沉积物DOC含量的测定水体叶绿素a测定参考《水质叶绿素a的测定分光光度法(HJ 897—2017)》.沉积物叶绿素a测定参考方法为:向沉积物中加入90%丙酮, 置于4 ℃萃取24 h, 用同步荧光法测定叶绿素含量(Hansson, 1988; 姜霞等, 2010).利用总有机碳分析仪(Shimadzu TOC-VCPH, Japan)测定沉积物提取液的DOC含量, 结果表示为沉积物DOC含量, 基本结果见表 1.
| 表 1 采样点位和样品基本信息 Table 1 Basic information of sampling sites, water and sediment samples |
利用紫外-可见分光光度计(UV-1800, Shimadzu)测量过滤上清液的紫外-可见吸收光谱, 测量波长范围为200~800 nm, 间隔为1 nm, 中速扫描, 以Milli-Q超纯水作为空白对照.采用如下公式计算和校正后的吸收系数(Shang et al., 2019; 周石磊等, 2019):
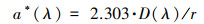
|
(1) |
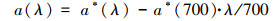
|
(2) |
式中, λ为波长(nm), a*(λ)和a(λ)分别为未经散射校正及已经散射校正后波长λ处的吸收系数(m-1);r为光程路径;D(λ)为波长为λ处的吸光度.本文选取a(254)来表示DOM的相对浓度.
DOM吸收光谱从紫外到可见区域随波长的增加呈现出指数衰减规律, 可用式(3)表示(Helms et al., 2008):
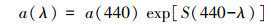
|
(3) |
式中, 440 nm为参照波长.S为指数函数曲线光谱斜率(μm-1).在本研究中, 分别对275~295 nm短波段和350~400 nm长波段进行拟合, 得到相应的光谱斜率S275~295和S350~400, 两者比值即为光谱斜率比值SR, 用来反映分子量或光降解等信息(Helms et al., 2008; Shang et al., 2019).
2.3.4 三维荧光光谱测定利用荧光分光光度计(F-7000, Hitachi)测定沉积物DOM三维荧光光谱.采用1 cm石英比色皿, 光源为150 W氙灯, 电压为700 V, 波长误差为±1 nm.其中, 波长的扫描范围为激发波长(Ex)从200~450 nm、发射波长(Em)从200~600 nm, 波长间隔均为5 nm, 扫描速度为12000 nm · min-1, 结果以Milli-Q超纯水作为空白.采用荧光区域积分(Fluorescence region integration, FRI)进行DOM组分分析, 根据荧光峰的位置不同, 可将三维荧光光谱划分为5个区域, 分别对应5类物质, 包括酪氨酸类蛋白质、色氨酸类蛋白质、类富里酸、溶解性微生物代谢产物和类腐殖酸.同时, 选用一系列荧光光谱指数来表征DOM性质, 包括荧光指数(Fluorescence index, FI)、腐殖化指数(Humification index, HIX)和生物源指数(Biological index, BIX), 这些参数的环境意义和计算方法见表 2.
| 表 2 区域荧光积分及荧光光谱指数 Table 2 Parameters of fluorescence region integration and fluorescence spectra index |
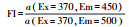
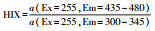

三维荧光光谱和各指标分布图由Origin 8.5制作.利用SPSS Statistics 18进行统计分析, 包括平均值、标准偏差的计算, 平均值t检验.
3 结果与讨论(Results and discussion) 3.1 紫外-可见吸收光谱特性及环境意义一般情况下, DOM紫外-可见光的吸收系数会随其含有化合物的芳香环缩合程度、含碳比例以及总含碳量的增加而增加(王晓江等, 2018).例如, 研究发现巢湖南淝河沉积物中溶解性有机碳与紫外-可见光吸收系数(a(254)、a(350)和a(440)等)呈现线性相关(p < 0.05), 说明紫外-可见光吸收系数a(λ)可以表征沉积物DOM的相对含量(沈烁等, 2014).巢湖表层四季沉积物DOM的a(254)均值为(32.2±16.2) m-1, 范围为13.1~101.7 m-1.在季节变化上, 巢湖春、夏、秋、冬四季表层沉积物中a(254)的平均值分别为(31.4±7.19)、(46.6±25.6)、(28.2±6.29)、(22.8±5.69) m-1 (图 2a).其中, 夏季a(254)含量要显著高于其它3个季节(p < 0.01), 这可能与夏季大规模爆发藻华产生较高生物量, 使得藻源溶解性有机质含量较高(姜霞等, 2010).从空间分布来看, 湖心区表层沉积物a(254)平均值和变异性((29.2±7.20) m-1)均小于北部湖滨区((35.8±22.4) m-1), 其中夏季湖滨区a(254)值超过了湖心区近2倍(图 2a).这可能与巢湖的季节常风向和藻的生长习性有关:在夏季和秋季, 漂浮在水面上的藻由于风浪作用最终在巢湖北部湖滨区集聚并沉降, 使得沉积物藻源性有机质呈现明显的时空差异(姜霞等, 2010; 殷长秦等, 2012).此外, 巢湖入湖支流众多, 湖滨区接收众多河流的废污水排放, 从而导致陆源输入的有机质在河口区累积, 使湖滨区表层沉积物a(254)值高于湖心区(王书航等, 2011; 李振涵等, 2018).
 |
| 图 2 巢湖沉积物DOM的紫外-可见光谱参数时空分布特征 (注:a、c和e分别表示不同季节表层沉积物DOM的a(254)含量、S275~295和SR值, 其中, 箱式图中间实线代表中位数, 方形代表均值;b、d和f分别表示夏季柱状样沉积物DOM的a(254)含量、S275~-295和SR值.) Fig. 2 Spatiotemporal distribution of ultraviolet-visible absorption spectrum index of DOM in sediments from Chaohu Lake |
巢湖沉积物湖心区(C4)和湖滨区(L5)两个沉积物柱状样a(254)值范围在10.1~42.6 m-1之间.从垂直分布上看, 随着深度的增加, a(254)先下降再增加, 两个样点均在10 cm处出现了极大值, 然后再逐渐下降, 直至深层趋于稳定(图 2b), 该趋势与陕西安康瀛湖沉积物中DOM的UV254(254 nm和365 nm处吸光度的差值)垂直分布特征一致(翟天恩等, 2017).河流中的悬浮颗粒物进入湖泊后因为水动力变缓会逐渐沉降在沉积物表面, 并与已有的沉积相、孔隙水和上覆水发生一系列物理、化学和生物作用, 其中的有机质也在这三相中保持动态平衡(沈吉等, 2010).由于有机质的矿化作用, 使得深层沉积物中的芳香族化合物含量明显低于表层沉积物层, 并逐渐趋于稳定.C4样点a(254)值大于L5样点, 这可能是由于湖心区受到多条入湖河流污染输入, 导致更多的有机质在此区域积累, 芳香族化合物的污染较重.而15 cm以下C4样点的a(254)值低于L5样点, 原因是湖心区C4样点深层沉积物受人类活动影响较少, 一些大分子质量DOM在长期的微生物作用下降解为小分子质量的组分(翟天恩等, 2017).
研究认为, S275~295能够反映沉积物中腐殖酸的变化情况, 即S值随着腐殖质中陆源组分与内源组分比值的减小而增大(沈烁等, 2014).本研究中, 巢湖表层沉积物DOM的S275~295均值为(25.3±5.60) μm-1, 范围为19.9~39.2 μm-1.在季节变化上, 春、夏、秋、冬四季表层沉积物的S275~295值分别为(21.4±1.1)、(22.3±1.4)、(23.2±1.98)和(34.1±3.40) μm-1(图 2c), 呈现明显的上升趋势, 且冬季S275~295要显著高于其它3个季节(p < 0.01), 这可能与富营养化湖泊中藻的生长循环对湖泊沉积物中DOM的影响有关.从春季到冬季, 富营养化湖泊中的藻类会经历“复苏-爆发-衰亡-休眠”的循环, 向湖泊沉积物中输入的藻源性有机质的量也逐步增加(姜霞等, 2010), 从而导致了陆源组分与自生源组分比值的减小(即S275~295值增加).在空间区域分布上, 湖心区表层沉积物S275~295((25.3±5.38) μm-1)与湖滨区((25.3±5.98) μm-1)无显著差异(图 2d), 这可能与巢湖西半湖区整体均处于严重富营养化水平有关(温胜芳, 2013).在垂直分布上, 随着深度的增加, 湖心区柱状样呈现稳定的缓慢上升, 但湖滨区柱状样波动剧烈, 且在12 cm处出现了极大值, 这可能是该样点处于湖滨带, 同时受到水华藻聚集沉降的藻源性有机质和陆源输入的有机质共同作用的影响.
光谱斜率比SR能很好地指示DOM中自生源所占的比重, SR越大, DOM中自生源组分含量就越多(Helms et al., 2008).巢湖表层沉积物SR均值为1.0±0.18, 范围为0.74~1.41(图 2e), 这与巢湖入湖河流南淝河沉积物SR范围基本一致(0.63~1.41) (沈烁等, 2014).其中夏季和冬季的SR较高, 说明在藻的爆发期和休眠期会向沉积物中输入较多的藻源性有机质使其自生源占DOM比例偏高.在空间分布上, 湖心区表层沉积物SR(1.02±0.19)与湖滨区(0.98±0.18)在统计上无显著差异(p > 0.05), 而且两点位柱状样的SR在垂直分布差异也不大(图 2f), 说明巢湖表层沉积物DOM特性受季节影响更大.
3.2 三维荧光的强度和组分特征巢湖沉积物DOM荧光强度呈明显的季节差异, 垂直分布上荧光强度总体呈下降趋势.夏季沉积物DOM荧光强度要显著高于其它3个季节(p < 0.05, 图 3a), 该结果与云南抚仙湖的季节性调查结果一致(陈乐等, 2019), 其可能的原因为:①夏季正值长江中下游流域(巢湖所在地)的雨季, 丰沛的降雨不仅能将陆源DOM通过入湖河流直接带入巢湖湖体, 同时也为湖泊中的微生物代谢提供相对充足的碳源(Zhou et al. 2015);②夏季是巢湖水华爆发最严重的时期, 相对较高的温度促使湖泊中微生物的新陈代谢活性增加, 从而向湖泊系统中输入大量藻源性有机质.基于十余年的野外监测也发现位于同一区域的太湖地区的太阳辐射和水温与叶绿素a呈显著正相关关系(p < 0.01), 即太阳辐射和水温的增加为湖泊中的水华爆发提供了良好的物理条件(陈桥等, 2009);③夏季巢湖水体表面会聚集大量的藻类, 导致阳光无法穿透表层水体, 从而有效地减少了DOM的光降解(Xu et al., 2013).此外, 夏季的湖心区表层沉积物DOM荧光强度要显著低于湖滨区(p < 0.01), 这可能与夏季藻华在风浪作用下最终聚集在北部的湖滨区有关(温胜芳2013).在垂直剖面上, 湖心区和湖滨区的荧光强度总体呈下降趋势(图 3b, 3c), 这可能是随着深度的增加, 营养性物质浓度的减少而导致微生物的丰度和活性减少(翟天恩等, 2017).
 |
| 图 3 巢湖沉积物DOM荧光强度和荧光组分比例 (a表示不同季节表层沉积物DOM荧光强度和组分;b和c分别表示L5和C4样点柱状沉积物DOM荧光强度和组分) Fig. 3 Relative intensities and proportions of DOM fluorescent components in sediments from Chaohu Lake |
结合区域荧光积分法(FRI)来计算出沉积物中DOM各个不同组分的相对比例.在季节分布上, 巢湖表层沉积物DOM中的PⅠ (酪氨酸类蛋白质)范围为8.3%~38.1%, 其中春季最高(32.7%±3.15%), 夏季最低(18.9%±4.54%), 秋冬两季维持在27%左右.DOM中的PⅡ(色氨酸类蛋白质)范围为12.4%~36.6%之间, 其中夏季最高(32.1%±3.46%), 秋冬季节相对较低(17.0%左右).DOM中的PⅢ(类富里酸)范围为6.75%~15.8%, 四季无明显差异, 秋季略高(12.1%±2.51%).DOM中的PⅣ (溶解性微生物代谢产物)范围为15.3%~46.3%, 春夏季较高(大于25%).DOM中的PⅤ (类腐殖酸)范围在8.24%~36.45%之间, 其分布规律与PⅣ正好相反, 春夏两季(小于15%)分别为低于秋冬两季(大于22%).巢湖沉积物DOM各组分平均占比的顺序为酪氨酸类蛋白质>溶解性微生物代谢产物>色氨酸类蛋白质>类腐殖酸>类富里酸(图 3a).类腐殖质一般是由地表径流携带的部分腐殖质以及浮游动植物释放的有机物经过细菌降解后产生, 而类蛋白主要由水生生物或微生物的新陈代谢作用产生(Rochellenewall et al., 2002).由此可见, 巢湖沉积物DOM组分分布受湖泊中藻生长循环的影响, 夏季由于温度较高, 微生物活动剧烈, 因此微生物代谢产物较多, 同时也带来了色氨酸类蛋白质组分的增加.巢湖沉积物DOM组分的垂直变化如图 3b、3c所示.随着深度的增加, 类腐殖酸、类富里酸和溶解性微生物代谢产物占比均不断下降.而沉积物DOM中类蛋白组分呈现波动式上升, 这表明近年来沉积物中DOM的组分趋于复杂化, 受多种来源影响(翟天恩等, 2017).
3.3 荧光光谱指数及其环境意义湖泊系统中DOM可分为陆源和生物源两种, 其中, 陆源DOM主要来自于流域土壤、地表径流、动植物残体以及人类活动输入的有机质, 以类腐殖质为主;而生物源DOM主要由藻类、水生生物、细菌等微生物降解和分泌物产生, 以类蛋白物质为主(Miller et al., 2010; Zhang et al., 2011).荧光光谱指数(FI、BIX和HIX)有效表征沉积物中DOM组分的来源, 结果见表 3.
| 表 3 巢湖沉积物DOM荧光光谱指数 Table 3 Fluorescence parameters for DOM samples in sediments from Chaohu Lake |
荧光指数(FI)常用来表征DOM中腐殖质的来源.一般认为, 当FI < 1.3时, DOM主要以径流输入为主, 水体和自身源贡献较少;而当FI > 1.9时, DOM主要来自微生物的代谢, 自生源特征突出(Fellman et al., 2010).本研究中, 巢湖表层沉积物FI范围为2.56~4.89, 均值为3.77, 并随着春夏秋冬时间变化呈现明显的上升趋势, 与S275~295变化趋势一致.湖心区和湖滨区的FI值无显著性的差异, 这可能是由于巢湖整个西半湖都处于相对严重的富营养状态, 从而没有表现出明显的空间差异.
腐殖化指数(HIX)是用来估算湖泊腐殖化程度的参数, 其值与DOM的腐殖化程度呈正相关.当HIX>6时, DOM具有强腐殖质特征, 且陆源贡献较大, 而当HIX<4时, 则表示DOM腐殖质特征较弱且以自生源为主, HIX若介于4~6之间则DOM属于强腐殖质特征和微弱的新近自生源(Ohno et al., 2007).本研究中, 巢湖表层沉积物DOM的HIX值均小于4, 这说明巢湖沉积物中DOM的腐殖质特征较弱并且以自生源为主.相较于其它季节, 秋、冬季HIX值分别为3.72±1.88和3.55±1.19, 要远高于春(0.76±0.15)、夏(1.38±0.44)两季, 说明秋冬季沉积物中DOM腐殖化程度更高, 稳定性更好, 这可能与秋冬季节温度较低, 微生物活性受到抑制有关.湖心区的HIX值(0.32±0.24)要低于湖滨区(0.61±0.10), 说明湖心区DOM自源性更高, 这可能是由于相对于湖滨区, 湖心区受到相对较小的外源干扰(Lei et al., 2016).
生物源指数(BIX)一般用于估计内源物质对水体中DOM的相对贡献以及DOM生物可利用性的高低.当BIX>1时, DOM主要是由细菌活动产生, 当BIX在0.8~1.0时, DOM具有较强的自生源性特征, 而当BIX < 0.6时, DOM主要来自于外源(卢松等, 2015).本研究中, 巢湖表层沉积物DOM中BIX值范围为0.31~1.54, 均值为0.87.秋冬季的BIX值相较于春夏季节较低, 特别是冬季的BIX均值<0.6, 这可能是由于秋冬季的温度较低, 从而导致微生物的活性减弱, 降低了其自源性DOM的生成.
结合我国不同水体沉积物DOM荧光光谱指数对比发现(图 4), 巢湖沉积物DOM中FI指数要显著高于其它水体, 如巢湖沉积物DOM平均FI指数(3.40)是水源地丹江口水库的2倍(1.71).HIX指数和BIX指数与其它水体范围基本一致, 但呈现出更大异质性, 这可能与本研究是4个季节采样, 而其它研究多为一次性采样有关.此外, 巢湖沉积物3个荧光指数指标的波动范围较大, 这说明巢湖沉积物DOM来源虽以自生源为主, 但具有陆源及内源的双重特征, 受藻类的生长循环影响较大.对巢湖主要入湖河流之一南淝河研究发现, 南淝河上游河流沉积物DOM更多来自陆地和土壤的陆源类腐殖质物质和类富里酸类物质, 而位于河流下游沉积物DOM多为自生源腐殖质, 这说明随着河流沿程变化, 沉积物DOM的组分和性质会呈现从陆源到自生源的转变(沈烁等, 2014; Zhang et al., 2020).后续研究应从流域层面出发, 考察“河流-河口-湖泊”水体和沉积物DOM组分和性质的转变, 重点关注湖泊系统中DOM组分的转变、内外源的贡献.
 |
| 图 4 我国不同水体沉积物DOM荧光光谱指数比较 (注:a、b和c分别表示荧光指数、生物指数和腐殖化指数.各代表水体基本介绍为:1.巢湖, 富营养化严重的西半湖, 数据来自本研究;2.蘑菇湖, 位于新疆石河子市, 现状水质为劣Ⅴ类(张广彩, 2019);3.蠡湖, 位于江苏省无锡市, 城市淡水景观湖泊(陈俊伊等, 2017);4.瀛湖, 位于陕西省安康市, 南水北调中线工程丹江口水库的重要水源(翟天恩等, 2017);5.周村水库, 位于山东省枣庄市, 城市供水水源地(周石磊等, 2019);6.金盆水库, 位于陕西省西安市, 黑河引水系统的主要水源地(王晓江等, 2018);7.丹江口水库, 位于湖北省十堰市和河南省南阳市, 南水北调中线工程水源地(冯可心等, 2014);8.南淝河, 位于安徽省合肥市, 巢湖的入湖河流, 水质多处于Ⅴ类或劣Ⅴ类(沈烁等, 2014);9.小浃江, 位于浙江省宁波市, 贯穿于城市的商业区、规模化工业区及部分农业区(高凤等, 2019);10.崇明岛, 位于上海市, 潮间带砂质钙质冲积型土壤(Wang et al., 2015).) Fig. 4 Comparison of fluorescence parameters for DOM in sediments from different water bodies in China |
富营养化是巢湖面临的主要水环境问题之一, 藻华爆发向湖泊中输入了大量藻源性有机质;与此同时, 巢湖地区社会经济的迅速发展, 流域内点源、面源污染形势依然严峻, 外源DOM输入也会间接促进内源DOM的增加.同时, 由于水体富营养化严重, 高的叶绿素含量会导致湖水透明度的降低, 从而削弱DOM的光化学降解, 进一步加剧了DOM丰度的提升.同时, 在光化学的作用下, 富营养化湖泊的藻源性有机质容易释放出被水生植物和微生物利用的小分子有机物质, 从而影响湖泊的富营养化进程(乔煜琦等, 2018).关注富营养化湖泊系统DOM的性质和组成, 对全面认识水生态系统中DOM调控污染物的生物地球化学循环过程具有重要意义.
4 结论(Conclusions)1) 巢湖表层沉积物夏季a(254)值要显著高于其它3个季节.垂直分布上, 随着深度的增加, a(254)值呈现先下降后增加的趋势, 在10 cm处出现峰值, 之后再逐渐下降直至深层并趋于稳定.表层沉积物DOM的S275~295值春夏秋冬呈现明显的上升趋势.
2) 巢湖沉积物DOM夏季的荧光强度要显著高于其它3个季节, 在垂直剖面上荧光强度总体呈下降趋势.DOM组分以酪氨酸类蛋白质和溶解性微生物产物为主, 各组分在空间分布上无明显差异.在垂直分布上, 各组分的强度整体呈现下降趋势.富营养化湖泊中藻的生长循环对湖泊沉积物DOM具有显著影响.
3) 巢湖沉积物DOM的荧光光谱指数中FI、HIX和BIX值的范围分别为2.56~4.89、0.57~6.78和0.31~1.54, 其季节差异较为显著, 而空间差异较小;在垂直分布上, 各指标整体呈下降趋势.3个指标均表明巢湖沉积物DOM主要来自于生物源, 但具有陆源及内源的双重特征.
Chen M, Jin H. 2015. Pre-treatments, characteristics, and biogeochemical dynamics of dissolved organic matter in sediments:A review[J]. Water Research, 79: 10-25. DOI:10.1016/j.watres.2015.04.018 |
Chen W, Westerhoff P, Leenheer J A, et al. 2003. Fluorescence excitation-emission matrix regional integration to quantify spectra for dissolved organic matter[J]. Environmental Science & Technology, 37(24): 5701-5710. |
陈俊伊, 王书航, 姜霞, 等. 2017. 蠡湖表层沉积物荧光溶解性有机质(FDOM)荧光光谱特征[J]. 环境科学, 38(1): 72-79. |
陈乐, 周永强, 周起超, 等. 2019. 抚仙湖有色可溶性有机物的来源组成与时空变化[J]. 湖泊科学, 31(5): 1357-1367. |
陈桥, 韩红娟, 翟水晶, 等. 2009. 太湖地区太阳辐射与水温的变化特征及其对叶绿素a的影响[J]. 环境科学学报, 29(1): 199-206. |
程红, 汪家权, 肖莆, 等. 2009. 巢湖流域农业非点源污染现状及控制策略[J]. 安徽农业科学, 37(29): 367-368+371. |
Dubois N, Saulnier-Talbot E, Mills K, et al. 2018. First human impacts and responses of aquatic systems:A review of palaeolimnological records from around the world[J]. Anthropocene Review, 5(1): 28-68. DOI:10.1177/2053019617740365 |
Fang T, Lu W, Li J, Zhao X, Yang K. 2017. Levels and risk assessment of metals in sediment and fish from Chaohu Lake, Anhui Province, China[J]. Environmental Science and Pollution Research, 24(11): 1-11. |
Fellman J B, Hood E, Spencer R G M. 2010. Fluorescence spectroscopy opens new windows into dissolved organic matter dynamics in freshwater ecosystems:A review[J]. Limnology & Oceanography, 55(6): 2452-2462. |
冯可心, 李永峰, 姜霞, 等. 2014. 丹江口水库表层沉积物有色可溶性有机物空间分布特征及其来源分析[J]. 环境化学, 35(2): 373-382. |
高凤, 邵美玲, 唐剑锋, 等. 2019. 城镇流域水体-沉积物中溶解性有机质的荧光特性及影响因素:以宁波市小浃江为例[J]. 环境科学, 40(9): 4009-4017. |
郭旭晶, 席北斗, 谢森, 等. 2012. 乌梁素海沉积物孔隙水中溶解有机质的荧光及紫外光谱研究[J]. 环境工程学报, 6(2): 440-444. |
Hansson L A. 1988. Chlorophyll a determination of periphyton on sediments:Identification of problems and recommendation of method[J]. Freshwater Biology, 20(3): 347-352. DOI:10.1111/j.1365-2427.1988.tb00460.x |
Helms J R, Stubbins A, Ritchie J D, et al. 2008. Absorption spectral slopes and slope ratios as indicators of molecular weight, source and photobleaching of chromophoric dissolved organic matter[J]. Limnology & Oceanography, 53(3): 955-969. |
Huang J, Zhang Y, Arhonditsis G B, et al. 2019. How successful are the restoration efforts of China's lakes and reservoirs?[J]. Environment International, 123: 96-103. DOI:10.1016/j.envint.2018.11.048 |
Huisman J, Codd G A, Paerl H W, et al. 2018. Cyanobacterial blooms[J]. Nature Reviews Microbiology, 16: 471-484. DOI:10.1038/s41579-018-0040-1 |
Huguet A, Vacher L, Relexans S, et al. 2009. Properties of fluorescent dissolved organic matter in the Gironde Estuary[J]. Organic Geochemistry, 40(6): 706-719. DOI:10.1016/j.orggeochem.2009.03.002 |
何伟, 白泽琳, 李一龙, 等. 2016. 水生生态系统中溶解性有机质表生行为与环境效应研究[J]. 中国科学:地球科学, 46(3): 93-107. |
姜霞, 王书航, 钟立香, 等. 2010. 巢湖藻类生物量季节性变化特征[J]. 环境科学, 31(9): 2056-2062. |
Lei P, Luís M N, Liu Y R, et al. 2019. Mechanisms of algal biomass input enhanced microbial Hg methylation in lake sediments[J]. Environment International, 126: 279-288. DOI:10.1016/j.envint.2019.02.043 |
Lei P, Zhang H, Shan B. 2016. Vertical records of sedimentary PAHs and their freely dissolved fractions in porewater profiles from the northern bays of Taihu Lake, Eastern China[J]. Rsc Advances, 6: 98835-98844. DOI:10.1039/C6RA11180G |
雷沛, 张洪, 王超, 等. 2018. 沉积物-水界面污染物迁移扩散的研究进展[J]. 湖泊科学, 30(6): 1489-1508. |
李振涵, 张文强, 雷沛, 等. 2018. 巢湖西部河口区沉积物氮磷分布特征与原位扩散通量估算[J]. 环境科学学报, 38(8): 2974-2982. |
卢松, 江韬, 张进忠, 等. 2015. 两个水库型湖泊中溶解性有机质三维荧光特征差异[J]. 中国环境科学, 35(2): 516-523. |
McKnight D M, Boyer E W, Westerhoff P K, et al. 2001. Spectrofluorometric characterization of dissolved organic matter for indication of precursor organic material and aromaticity[J]. Limnology & Oceanography, 46(1): 38-48. |
Miller M P, Mcknight D M. 2010. Comparison of seasonal changes in fluorescent dissolved organic matter among aquatic lake and stream sites in the Green Lakes Valley[J]. Journal of Geophysical Research:Biogeosciences, 115(12): 1-12. |
Ohno T, Fernandez I J, Hiradate S, et al. 2007. Effects of soil acidification and forest type on water soluble soil organic matter properties[J]. Geoderma, 140(1/2): 176-187. |
Pitta E, Zeri C, Tzortziou M, et al. 2017. Seasonal variations in dissolved organic matter composition using absorbance and fluorescence spectroscopy in the Dardanelles Straits-North Aegean Sea mixing zone[J]. Continental Shelf Research, 149: 82-95. DOI:10.1016/j.csr.2016.07.013 |
乔煜琦, 江海洋, 李星, 等. 2018. 蓝藻水华暴发和衰亡对太湖有色可溶性有机物的影响[J]. 湖泊科学, 30(4): 907-915. |
Rochellenewall E J, Fisher T R. 2002. Production of chromophoric dissolved organic matter fluorescence in marine and estuarine environments:An investigation into the role of phytoplankton[J]. Marine Chemistry, 77(1): 7-21. |
Shang Y, Song K, Jacinthe P A, et al. 2019. Characterization of CDOM in reservoirs and its linkage to trophic status assessment across China using spectroscopic analysis[J]. Journal of Hydrology, 576: 1-11. DOI:10.1016/j.jhydrol.2019.06.028 |
尚丽霞, 柯凡, 李文朝, 等. 2013. 高密度蓝藻厌氧分解过程与污染物释放实验研究[J]. 湖泊科学, 25(1): 47-54. DOI:10.3969/j.issn.1003-5427.2013.01.007 |
沈吉, 薛滨, 吴敬禄. 2010. 湖泊沉积与环境演化[M]. 北京: 科学出版社.
|
沈烁, 王育来, 杨长明, 等. 2014. 南淝河不同排口表层沉积物DOM光谱特征[J]. 中国环境科学, 34(9): 2351-2361. |
孙共献. 2015.水华微囊藻越冬、春季复苏机制及衰亡后营养盐释放过程研究[D].安徽: 安徽师范大学 http://d.old.wanfangdata.com.cn/Thesis/D744525
|
Villacorte L O, Ekowati Y, Neu T R, et al. 2015. Characterisation of algal organic matter produced by bloom-forming marine and freshwater algae[J]. Water Research, 73: 216-30. DOI:10.1016/j.watres.2015.01.028 |
Wang Y L, Yang C M, Zou L M, et al. 2015. Spatial distribution and fluorescence properties of soil dissolved organic carbon across a riparian buffer wetland in Chongming Island, China[J]. Pedosphere, 25(2): 220-229. DOI:10.1016/S1002-0160(15)60007-8 |
王书航, 姜霞, 金相灿. 2011. 巢湖入湖河流分类及污染特征分析[J]. 环境科学, 32(10): 2934-2839. |
王晓江, 黄廷林, 李楠, 等. 2018. 峡谷分层型水源水库表层沉积物溶解性有机物光谱特征[J]. 湖泊科学, 30(6): 1625-1635. |
温胜芳. 2013.一种沉积物结构研究的方法学构建及在巢湖的应用研究[D].北京: 中国科学院大学 http://d.wanfangdata.com.cn/thesis/Y2369032
|
吴丰昌. 2010. 天然有机质及其与污染物的相互作用[M]. 北京: 科学出版社.
|
Xu H, Jiang H. 2013. UV-induced photochemical heterogeneity of dissolved and attached organic matter associated with cyanobacterial blooms in a eutrophic freshwater lake[J]. Water Research, 47(17): 6506-6515. DOI:10.1016/j.watres.2013.08.021 |
殷长秦, 王体健, 石春娥, 等. 2012. 巢湖流域典型站点的风场特征分析[J]. 大气科学学报, 35(2): 123-130. |
Zhang H, Cui K P, Guo Z, et al. 2020. Spatiotemporal variations of spectral characteristics of dissolved organic matter in river flowing into a key drinking water source in China[J]. Science of the Total Environment, 700: 134360. DOI:10.1016/j.scitotenv.2019.134360 |
Zhang Y L, Dijk M A V, Liu M L, et al. 2009. The contribution of phytoplankton degradation to chromophoric dissolved organic matter (CDOM) in eutrophic shallow lakes:field and experimental evidence[J]. Water Research, 43(18): 4685-4697. DOI:10.1016/j.watres.2009.07.024 |
Zhang Y, Yin Y, Feng L, et al. 2011. Characterizing chromophoric dissolved organic matter in Lake Tianmuhu and its catchment basin using excitation-emission matrix fluorescence and parallel factor analysis[J]. Water Research, 45(16): 5110-5122. DOI:10.1016/j.watres.2011.07.014 |
Zhang Y L, Zhou Y Q, Shi K, et al. 2018. Optical properties and composition changes in chromophoric dissolved organic matter along trophic gradients:Implications for monitoring and assessing lake eutrophication[J]. Water Research, 131: 255-263. DOI:10.1016/j.watres.2017.12.051 |
赵萱, 成杰民, 鲁成秀. 2012. 不同生态类型富营养化湖泊沉积物中有机质赋存形态[J]. 环境化学, 31(3): 302-307. |
Zhou Y, Zhang Y, Shi K, et al. 2015. Dynamics of chromophoric dissolved organic matter influenced by hydrological conditions in a large, shallow, and eutrophic lake in China[J]. Environmental Science and Pollution Research, 22(17): 12992-13003. DOI:10.1007/s11356-015-4556-x |
Ziegelgruber K L, Zeng T, Arnold W A, et al. 2013. Sources and composition of sediment pore-water dissolved organic matter in prairie pothole lakes[J]. Limnology & Oceanography, 58(3): 1136-1146. |
Zsolnay A, Baigar E, Jimenez M, et al. 1999. Differentiating with fluorescence spectroscopy the sources of dissolved organic matter in soils subjected to drying[J]. Chemosphere, 38(1): 45-50. DOI:10.1016/S0045-6535(98)00166-0 |
翟天恩, 霍守亮, 张靖天. 2017. 沉积物中溶解性有机质的垂直分布光谱特性[J]. 环境工程学报, 11(11): 416-424. |
张广彩. 2019.蘑菇湖沉积物中有机质光谱指纹特征与迁移转化机制研究[D].济南: 山东师范大学
|
张民, 孔繁翔. 2015. 巢湖富营养化的历程、空间分布与治理策略(1984-2013)[J]. 湖泊科学, 27(5): 791-798. |
朱广伟, 许海, 朱梦圆, 等. 2019. 三十年来长江中下游湖泊富营养化状况变迁及其影响因素[J]. 湖泊科学, 31(6): 1510-1524. |
周石磊, 孙悦, 张艺冉, 等. 2019. 山东省周村水库季节演替中沉积物上覆水溶解性有机物的紫外-可见与三维荧光光学特征[J]. 湖泊科学, 31(5): 1344-1356. |