 2020, Vol. 40
2020, Vol. 40



2. 北京林业大学林学院, 北京 100083
2. College of Forest Science, Beijing Forestry University, Beijing 100083
土壤作为陆地生态系统最大的碳库, 其碳库储量的增加对于减缓气候变化进程具有重要意义(Lal, 2004).土壤有机碳(Soil organic carbon, SOC)储量主要取决于碳源输入与输出之间的平衡(Schippers et al., 2004).大气CO2浓度的持续增加引起的“CO2施肥效应”通过影响植被光合作用及其总初级生产力, 改变凋落物的数量和土壤碳的输入(Schippers et al., 2004), 而后者的变化又能够通过影响土壤的生物化学过程影响土壤碳的输出(Davidson et al., 2006).氮是陆地生态系统尤其是温带森林中常见的限制性养分, 大气氮沉降的增加可在一定程度上减轻植被和土壤的氮受限程度, 促进植物生长及增加或降低不同质量凋落物分解速率(Norby et al., 2010), 也能调控土壤微生物底物利用效率及其生长代谢, 或抑制微生物酶的活性而增加土壤碳的储量(Frey et al., 2004), 或促进碳降解相关胞外酶的释放而增加土壤有机碳的分解.我国作为全球氮沉降三大热点地区之一, 从1980—2010年全国大气氮沉降量平均值由11.11 kg · hm-2 · a-1增加到13.87 kg · hm-2 · a-1(Jia et al., 2014), 但由于氮沉降量分布的时空差异性及土壤类型的多样性和复杂性, 科学评估大气氮沉降对土壤碳动态的影响及其生态效应仍然是目前研究的热点和盲点(Tian et al., 2015).例如, 土壤不同物理(团聚体结构)和化学组成的碳组分对气候变化的响应不同, 长期施氮和地上碳源输入变化之间的交互作用对不同碳、氮组分的影响更是存在很大的不确定性(Sun et al., 2016; Sun et al., 2019).因此, 探究土壤不同碳、氮组分对土壤碳、氮输入变化的响应, 对于预测未来土壤生物地球化学元素动态具有重要意义.
植物凋落物作为土壤碳的主要来源, 经过土壤微生物和酶等生物有机体的破碎、分解转化后, 其残茬和微生物代谢产物以可溶性的化合物等形式淋溶进入土壤, 最终以不同的物理化学形态赋存于土壤中(Schmidt et al., 2011; Cotrufo et al., 2013).然而, 不同数量和质量的凋落物由于其分解难易程度和速率不同, 转化进入和固持在土壤中的量不同(Berti et al., 2016).例如, 相对于输入速率, 不稳定的植物源碳由于其更容易被微生物有效利用, 成为微生物代谢的主要能量来源(Tian et al., 2016).这些分解后的微生物产物作为相对稳定的土壤有机碳的主要前体(Dungait et al., 2012), 通过与矿物土基质的强化学键结合, 促进土壤团聚体的形成, 从而降低微生物等分解者的可接触性, 进一步增强了土壤碳的相对稳定性(Dungait et al., 2012).团聚体作为土壤的重要结构组成, 参与并调控了土壤的生物物理化学过程, 尤其是作为土壤碳贮存的载体, 对于碳的固持具有重要意义(Dungait et al., 2012).然而, 土壤碳氮组分对气候变化响应的差异, 很大程度上可能是由于土壤底物C : N化学计量组成的不同, 资源分配的差异促使微生物通过调整自身C : N化学计量或分泌不同的酶等选择性代谢底物能量和养分(Allison et al., 2005).因此, 对于含氮量有限的温带森林, 在改变土壤碳源输入条件下增加氮的输入能够进一步揭示土壤碳、氮响应土壤底物浓度变化的关键机制.
土壤碳、氮组分的动态是其不同物理、化学组分形态结构交互变化的综合过程, 笼统地将土壤碳、氮作为单一整体进行研究可能会忽视土壤碳、氮转化过程中的关键环节.因此, 本研究基于长期野外观测样地建立的地上凋落物处理和氮添加双因素随机区组实验, 分析森林表层土壤碳、氮组分及其化学计量比的响应特征, 并提出以下假设:①长期地上凋落物输入量的加倍增加了碳源的输入而显著促进了土壤有机碳的含量, 同时高质量凋落物由于其较快的分解速率, 相对于低质量的枝果凋落物将显著增加土壤有机碳的含量;②氮元素的添加显著增加了微生物等分解者的养分有效性, 促进了凋落物的分解和微生物碳源的代谢利用(Wang et al., 2017), 从而降低了土壤活性碳氮组分的含量;③土壤大团聚体相较于黏粉粒碳氮组分更易受外界环境变化的影响, 因此, 地上凋落物和氮添加处理将显著影响大团聚体碳氮组分的含量;④由于土壤微生物碳源(能量)和氮源(养分)输入量的增加, 将显著促进微生物的生物量碳、氮大小.
2 材料与方法(Materials and methods) 2.1 研究区概况研究区位于山西太岳山林管局灵空山林场(36°33′28″~36°42′52″N, 111°59′27″~112°07′48″E), 属暖温带半干旱大陆季风气候, 地势北高南低, 海拔为1450~1600 m, 年均温为8.6 ℃, 年均降水为662 mm, 集中于7、8、9月(涂玉等, 2012).土壤主要为石灰岩母岩发育而成的山地褐土、山地淋溶褐土和山地棕壤, 本样地土壤pH为6.5, 平均总有机碳(Total organic carbon, TOC)、全氮含量(Total nitrogen, TN)和土壤碳氮比值分别为73.8 g · kg-1、4.7 g · kg-1和13.6.研究区植被区划属于北暖温带落叶阔叶林地带, 主要乔木树种为油松(Pinus tabuliformis)、辽东栎(Quercus liaotungesis)、山杨(Populus davidiana)和白桦(Betula platyphylla)等.
2.2 样地设置本研究设置不同数量、质量的地上凋落物处理和不同浓度的氮添加, 建立随机区组实验, 具体为:在样地内布设60个2 m×2 m的小样方, 共15列4行, 每3列划分为1个区组, 缓冲间隔1 m, 小样方间距0.5 m;各区组内, 分别完全随机设置3个浓度的氮添加和4种凋落物添加的交互处理, 每列进行相同的氮添加量处理.其中, 地表凋落物处理包括凋落物剔除(Lnil)、木质(枝果)凋落物加倍(Lwoody)、叶凋落物加倍(Lleaf)和枝果叶混合凋落物加倍(Lmix)共4个处理, 具体处理方法为:将原先位于Lnil样方腐殖质层及样方中的树枝(包括细碎小枝)、果实和叶凋落物挑拣出来后分别添加到Lwoody和Lleaf样方中, 将Lwoody样方中的叶片和Lleaf样方中的枝果挑拣出来添加到Lmix样方中;氮添加处理设置3个水平, 分别为0 (N0)、5 (N5)和10 g · m-2 · a-1(N10).施用氮肥为尿素, 于2009年9月份建立实验样地后, 每年4、6、8、10月按照既定实验设计进行氮添加和凋落物处理及样地维护.本研究样地建于该地区具有典型代表的油松-辽东栎天然次生林内, 样地平均叶、枝和果(主要为松球和辽东栎种子)凋落物生物量分别约为366、158和253 g · m-2 · a-1.
2.3 样品采集与测定土壤样品采集于2017年8月, 在样地每个小样方内小心拨开地表枯枝落叶层, 选取同一小样方内中心点及4个方位角位置, 分别取0~5 cm表层土, 将采集到的5个土样混合, 剔除石砾、根系及动植物残体, 四分法取样后带回实验室, 置于室内阴凉处自然风干, 用于土壤理化性质分析(鲍士旦等, 1988).其中, 土壤含水量采用烘干称重法测定;土壤容重(Bulk density, BD)采用环刀法测定;土壤pH值用奥豪斯pH计测定(水土比2.5 : 1);土壤阳离子交换量(Cation exchange capacity, CEC)采用乙酸铵法测定;土壤总有机碳含量(Total organic carbon, TOC)采用重铬酸钾-浓硫酸高温外加热氧化法测定;土壤全氮含量(Total nitrogen, TN)采用凯氏定氮仪测定;土壤铵态氮(NH4+-N)和硝态氮(NO3--N)采用连续流动分析仪测定;土壤活性碳组分含量(Labile organic carbon, LOC)采用高锰酸钾氧化法测定(Blair et al. 1995), 其碳组分占比计算为fLOC=(LOC/TOC)×100%;土壤芳香度指数利用分光光度计比色测定, 具体为:对土壤样品进行浸提, 然后用石英比色皿在254 nm波长下进行扫描, 利用公式(1)计算SUVA254(L · mg · m-1), 以表征土壤可溶性有机物质(Dissolved organic matter, DOM)芳香性大小(Jaffrain et al. 2007).
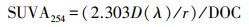
|
(1) |
式中, D(λ)为吸光度, r为光程路径(m), DOC(Dissolved organic carbon)为土壤样品浸提后的可溶性有机碳的含量(mg · L-1), SUVA254值越大, 表明DOM芳香性越强.
土壤团聚体的分离采用湿筛法(Six et al., 2000), 具体为通过土壤团聚体分析仪(XY-100型, 北京祥宇)中不同孔径的土壤套筛置于一定体积的蒸馏水中上下振荡(40次· min-1, 振幅4 cm) 10 min, 得到大团聚体(Macro-aggregation, 250~2000 μm)、微团聚体(Micro-aggregation, 53~250 μm);留在蒸馏水中的土壤颗粒为土壤黏粉粒(< 53 μm), 采用离心法(TGL-16M, 湖南湘仪)分离出黏粒(Silt-aggregation, 2~53 μm, 1300 r · min-1, 7 min)和粉粒(Clay-aggregation, < 2 μm, CaCl2溶液凝絮, 4800 r · min-1, 15 min).因本样地土壤为山地棕壤, 湿筛后并未有效获取大于2000 μm的组分.之后将各粒径土壤团聚体置于烘箱中50 ℃烘干称重并计算质量百分比, 分别为WMacro、WMicro、WSilt、WClay.土壤团聚体稳定性的指标计算采用平均质量直径(Mean weight diameter, MWD, mm), 值越大则土壤团聚体结构越稳定(Le Bissonnais et al. 2018):
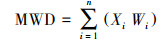
|
(2) |
式中, Xi和Wi分别为不同粒径大小团聚体的平均直径(mm)和质量百分比.
2.4 数据处理本研究野外样地设计采用的是随机区组实验设计, 为了量化地上凋落物处理和氮添加对土壤碳氮组分的影响, 采用线性混合效应模型分别对不同有机碳组分、全氮含量及各组分的碳氮比值等进行了分析计算.其中, 将地上凋落物处理(4个处理)和施氮量(3个水平)及这两个处理间的交互作用作为固定因素, 将各区组作为随机因素处理, 并采用LSD检验(Least significant difference test)对不同处理进行事后多重比较.在分析之前, 首先对各变量数据进行正态性检验, 对不符合正态分布的变量进行数据转换.采用R语言中“lmerTest”软件包进行线性混合模型分析, 显著性水平p值设为0.05.同时, 采用Pearson相关系数计算土壤不同有机碳、氮组分及各组分C : N比值与土壤环境因子之间的相关性.
3 结果与分析(Results and analysis) 3.1 土壤理化性质和团聚体粒径组成响应特征经过8年的地上凋落物处理和氮添加实验, 土壤pH、CEC、NH4+-N、NO3--N对两个处理的响应均未表现出显著性差异(p>0.05), 但长期地上凋落物处理显著影响了土壤水分含量和容重, 具体表现为:Lmix处理相较于Lcon和Lnil处理显著增加了土壤含水量, 而Lmix和Lleaf处理显著降低了土壤容重;氮添加显著降低了土壤容重, 对土壤其他理化属性无显著性影响.同时, 不同团聚体质量组成对地上凋落物处理和氮添加响应存在差异:含叶凋落物加倍输入(Lmix和Lleaf)相较于凋落物去除处理(Lnil)显著促进了土壤大团聚体质量百分比, 并增加了土壤团聚体的稳定性指标(MWD)(表 1);与此相反的是, Lnil处理显著增加了团聚体黏粒组分(Silt)质量百分比, 且枝果凋落物加倍处理(Lwoody)显著增加了粉粒组分(Clay)质量百分比;氮添加并没有显著影响土壤团聚体质量的分布(表 1).
| 表 1 土壤团聚体质量百分比及其平均质量直径对地上凋落物处理和氮添加的响应特征(均值±标准误) Table 1 The response characters of the percentage by weight of soil aggregate sizes and its mean weight diameter to forest aboveground litter manipulation and nitrogen addition (Mean ± SE) |
除了可溶性碳氮比值之外, 地上凋落物处理和氮添加显著影响了土壤各碳、氮组分的含量及其芳香性大小(SUVA254), 但两个处理均没有表现出显著的交互作用(图 1).具体表现为:地上含叶凋落物加倍处理(Lleaf)相较于Lnil处理显著增加了土壤总有机碳(TOC)、全氮(TN)、碳氮比值(C : N)、可溶性碳(DOC)、活性碳(LOC)的含量;Lnil、Lwoody处理相较于Lleaf、Lmix处理显著增加了SUVA254值;凋落物处理中的Lleaf和Lmix对各指标并没有表现出显著性差异, 且仅Lcon处理中的DON含量显著低于Lleaf和Lmix处理, 其他各变量中Lcon处理相较于Lleaf和Lmix处理并无显著性差异(图 1).氮添加处理中N5相较于N0处理显著促进了TOC、TN、DON、LOC含量和fLOC值;N5与N10处理对各变量的影响并无显著性差异, 仅变量LOC和fLOC在N5处理下显著高于N10和N0处理.
 |
| 图 1 土壤不同碳、氮组分及其比值、活性碳占比和芳香性对地上凋落物处理和氮添加的响应特征(均值±标准误) Fig. 1 The response characters of soil carbon and nitrogen fractions, and their C : N ratios, the percentage of labile organic carbon and SUVA254 to forest aboveground litter manipulation and nitrogen addition (Mean ± SE) |
地上凋落物处理显著影响了土壤团聚体碳、氮含量, 但仅有大团聚体(250~2000 μm)和微团聚体(53~250 μm)组分碳、氮含量对地上凋落物处理和氮添加的响应达到统计学显著性水平, 且4个团聚体组分碳氮比值均未对两个处理呈现显著的交互作用(图 2).其中, 含叶凋落物处理(Lleaf和Lmix)相较于Lcon、Lnil和Lwoody处理均显著促进了土壤大团聚体和微团聚体碳、氮(Macro-C/N和Micro-C/N)的累积;对于不同团聚体碳氮比值, Lleaf、Lmix处理相较于Lcon、Lnil和Lwoody处理仅显著增加了微团聚体碳氮比值大小(Micro-C : N ratio);氮添加处理中, N5相较于N0仅显著促进了大团聚体有机碳和全氮含量, 对其他团聚体碳、氮含量并无显著性影响, 且N10与N5处理对4个团聚体碳、氮含量和比值的影响并无显著性差异.尽管土壤大、微团聚体碳、氮含量对地上凋落物处理和氮添加响应显著, 但不同团聚体碳、氮相对含量在两个处理条件下均没有显著性差异(p>0.05).地上凋落物处理中Lmix和Lleaf增加了土壤微生物生物量碳(MBC)的含量, 但并未达到统计学上的显著性水平(p=0.09);表层土壤微生物生物量氮(MBN)及其与MBC的比值(MBC : MBN)也并未因地上凋落物处理和氮添加而产生显著性差异(图 3).
 |
| 图 2 土壤团聚体碳、氮及其比值对地上凋落物处理和氮添加的响应特征 (均值±标准误) Fig. 2 The response characters of soil aggregate organic carbon and nitrogen and their ratios to forest aboveground litter manipulation and nitrogen addition (Mean ± SE) |
 |
| 图 3 土壤微生物生物量碳、氮及其比值对地上凋落物处理和氮添加的响应特征 (均值±标准误) Fig. 3 The response characters of soil microbial biomass carbon and nitrogen and their ratios to forest aboveground litter manipulation and nitrogen addition (Mean±SE) |
Pearson相关性分析表明(图 4), 土壤理化性质中, pH与土壤团聚体碳、氮组分并无显著相关关系, 土壤容重(BD)与不同团聚体碳、氮含量整体成负相关关系, 其他各指标均成正相关关系, 且BD和土壤质量含水量(SWC)整体与各组分碳、氮含量均显著相关.微生物生物量碳、氮也与土壤各碳、氮组分含量成正相关关系, 但MBC : MBN与各组分碳、氮含量并未有显著相关性.不同组分碳、氮含量与其碳氮比值亦成正相关关系, 其中, TOC与各组分碳氮比值均达到显著性关系.与土壤不同碳、氮组分绝对含量不同的是, 各组分碳、氮组分相对含量与土壤理化属性和碳氮比值更多地呈现负相关关系, 比如:大、黏粒团聚体碳、氮百分比含量(fMacro-C/N、fClay- C/N)及各团聚体质量百分比含量(WMicro、WSilt、WClay).土壤平均质量直径(MWD)与各碳、氮组分含量及其碳氮比均成正相关关系, 与土壤理化性质中阳离子交换量(CEC)成显著正相关关系.
 |
| 图 4 土壤不同碳、氮组分属性与土壤理化性质和微生物生物量碳、氮属性之间Pearson相关系数矩阵 (注:BD:容重;SWC:土壤质量含水量;CEC:阳离子交换量;MWD:团聚体平均质量直径;Macro-C : N、Micro-C : N、Silt-C : N、Clay-C : N:大、微和黏粉粒团聚体组分碳氮比;TOC:总有机碳;TN:全氮;DOC、DON:可溶性碳、氮;Macro-C/N、Micro-C/N、Silt-C/N、Clay-C/N:大、微和黏粉粒团聚体组分碳或氮含量;fMacro-C/N、fMicro-C/N、fSilt-C/N、fClay-C/N:大、微和黏粉粒团聚体组分碳或氮含量百分比) Fig. 4 The correlation coefficient matrix between soil carbon and nitrogen fractions, soil environment factors and soil microbial biomass carbon and nitrogen |
温带森林土壤有机碳占全球森林有机碳储量的13.77%(Pan et al., 2011), 对全球变化因素响应敏感, 如气候变化引起的碳输入和输出变化, 以及氮沉降引起的土壤养分有效性变化等.尽管许多研究已经探讨了全球变化对温带森林土壤有机碳动态的影响(Frey et al., 2018; Forstner et al., 2019), 但不同实验设计和实验持续周期的结果表明土壤碳氮对于气候变化的响应仍存在极大的不确定性(Xu et al., 2013; Lajtha et al., 2018).Meta分析的结果也表明土壤碳氮组分及其化学计量在CO2浓度升高和N添加分别处理条件下仍然存在较大的变异性(Yang et al., 2011).其中, 一个关键的问题即:全球变化因素如何调节SOC的组成及微生物的响应特征, 这个不确定性限制了对全球变化下长期SOC动态的理解.通过对温带森林地上凋落物8年的野外样地处理和氮添加实验, 本研究的结果表明土壤碳源和氮源输入的变化对土壤各有机碳、氮组分和微生物生物量碳、氮均有显著影响.
4.1 地上凋落物处理和氮添加对土壤不同碳、氮组分的影响地上凋落物的输入是土壤有机碳的重要来源, 地上生物量和凋落物量与土壤有机碳含量成显著正相关关系(Belay-Tedla et al., 2009).本研究中, 地上含叶凋落物处理(Lleaf和Lmix)相较于凋落物去除处理(Lnil)均显著促进了土壤总有机碳、可溶性碳、活性碳及大、微团聚体有机碳组分的含量(图 1、图 2), 即地上碳源加倍输入促进了土壤有机碳的累积, 与本研究假设1相符.尽管枝果凋落物也进行了加倍处理(Lwoody), 但与对照和凋落物去除相比其对土壤各碳组分的影响并无显著性差异(图 1、图 2), 这可能是由于枝果凋落物相较于含叶凋落物, 其本身分解速率较慢, 不足以在本实验处理周期内显著影响土壤有机碳组分的含量.同时, 微生物对碳源的选择性利用, 优先选择高质量易分解的凋落物, 这也间接造成含叶凋落物输入对土壤碳组分影响的显著效应.有研究通过对经同位素标记的凋落物进行分解实验, 结果表明, 叶和根凋落物分解过程中产生的微生物化合物对矿质土壤贡献了大量的碳, 而含木质素和酚类等相对难降解的植物组分不会优先积累在土壤有机质中(Mambelli et al., 2011).另外一个值得注意的现象是, 凋落物去除和加倍处理与对照相比, 整体上并没有显著降低或增加土壤碳组分的含量, 尤其是凋落物去除处理对所有的碳组分与对照相比均无显著性差异(图 1、图 2), 这可能是由于本实验仅对地上凋落物输入进行了处理,并没有对地下根系碳源的输入设置对照.大量研究表明, 地下根系碳源的输入相较于地上凋落物的输入, 其分解缓慢且根系形成的碳相对滞留时间更长, 对土壤碳的含量贡献更大(Sokol et al., 2019).氮沉降显著影响着土壤的碳循环, 本研究中地上凋落物处理和氮添加两者之间对不同土壤碳、氮组分均未表现出显著的交互作用(图 1~3), 但氮添加显著促进了土壤全氮、可溶性氮和大团聚体氮的含量(图 1、图 2), 表明氮沉降促进了土壤各氮组分的累积;同时, 氮添加显著促进了土壤总有机碳、活性碳组分和大团聚体碳的含量(图 1、图 2), 这与本研究假设2并不相符, 这可能是由于氮添加抑制了土壤碳循环的过程, 如土壤呼吸、土壤有机碳分解转化过程中酶的活性, 并诱导了难降解性有机碳的积累(Silva-Sánchez et al., 2019).
此外, Lleaf和Lmix处理相较于Lnil和Lwoody处理显著降低了土壤可溶性有机物质的芳香性(SUVA254)(图 1), 土壤有机物芳香性的改变是土壤微生物等生物有机体代谢碳源过程中产生的不同化学组分有机物质的结果(Näthe et al., 2017), 因此, 高质量凋落物的添加降低了土壤SUVA254.这可能是由于碳源和氮源的增加显著刺激了微生物的代谢活性并优先利用有机酸等碳组分来改变土壤DOM的数量和生物降解性(Dieckow et al., 2005), MBC与SUVA254成显著负相关(p < 0.05)及本研究样地的前期实验也表明Lleaf和Lmix处理显著促进了微生物的活性(Sun et al., 2016), 均进一步说明了微生物在土壤碳源转化中的主导作用.微生物作为土壤有机质分解转化的“引擎”, 对外界缓解变化敏感.然而, 本研究中微生物生物量碳、氮及其比值均对地上凋落物处理和氮添加响应不显著, 仅MBC达到边际显著水平(p < 0.1), 这与本文假设4并不相符, 这可能是微生物对外界环境长期变化适应的结果(Wallenstein et al., 2012).同时, 本次微生物属性分析只是单独一次采样的结果, 并不能完全表征长期条件下土壤微生物的响应特征.土壤有机碳和氮的动态变化是微生物等生物有机体分解代谢和利用的过程(朱仁欢等, 2018), 未来关于土壤碳氮的动态, 应更多地关注土壤微生物的时空动态响应特征及与土壤碳氮组分之间的协调机制, 以便更准确地揭示未来气候变化条件下土壤碳、氮的响应特征.
4.2 地上凋落物处理和氮添加对土壤团聚体属性的影响本研究中, 地上凋落物处理显著影响了土壤大、微团聚体碳、氮组分含量, 但土壤黏粉粒组分碳、氮含量响应微弱(图 2), 与本研究假设3相符.这是由于土壤团聚体本身的周转过程缓慢, 土壤黏粉粒组分相对更加稳定, 不易受外界环境变化的影响(Six et al., 2000).同时, 根据土壤团聚体的形成机制, 凋落物的添加可促进有机碳的积累, 其作为胶结物质可促进土壤团聚体的形成, 进而形成大团聚体, 且有机碳优先在大团聚体中累积, 在大团聚体内部逐渐破碎并结合到“微”团聚体(< 53 μm)中, 而后与微团聚体一起释放出来形成稳定的不易分解的碳组分(Vogel et al., 2014), 氮添加也促进了这一过程(Janssens et al., 2010).土壤团聚体的稳定性直接关系到土壤碳、氮组分的稳定性, 越来越多的观点认为土壤碳的累积和稳定并不是由于有机质本身的难降解性, 而是由于土壤团聚体的吸附包裹阻断了微生物等生物有机体的直接利用(Dungait et al., 2012).本研究中, 土壤各碳、氮组分与团聚体稳定性指标(MWD)成正相关关系(图 4), 地上含叶凋落物的加倍输入(Lleaf和Lmix)也显著促进了MWD(表 1), 这可能是由于碳源输入在促进土壤团聚体形成的同时增加了土壤碳、氮组分的含量(Bucka et al., 2019).土壤阳离子交换量(CEC)与MWD成显著正相关(p < 0.01), 与各碳、氮组分均成正相关关系(图 4), 可能也是通过促进土壤团聚体通过形成有机-矿物复合物来增加土壤团聚体及其碳、氮组分稳定性的途径之一(Le Bissonnais et al., 2018).这是由于土壤金属氧化物主要以铁铝氧化物的形式和粘土矿物通过配位体置换、高价离子键桥等作用降低土壤有机碳组分的生物有效性, 从而增加其稳定性和土壤的固持(von Lutzow et al., 2006).从土壤团聚体角度探究土壤碳、氮组分对外界环境变化的响应特征, 为观测土壤碳氮动态提供了重要视角, 对未来进一步揭示土壤不同尺度粒径中微生物和酶的响应特征奠定了基础.
5 结论(Conclusions)长期地上叶凋落物输入处理和氮添加显著促进了土壤不同物理(团聚体)和化学碳氮组分的含量, 但由于土壤黏粉粒团聚体周转缓慢, 在本实验周期内其碳、氮组分含量并未受到地上碳源和氮源处理的显著影响;土壤总的C : N比值与微团聚体(53~250 μm)C : N比值对地上高质量(低C : N比)碳源输入响应显著, 氮添加并没有显著改变土壤各组分的C : N比值.土壤微团聚体在促进和维持土壤碳、氮含量及团聚体稳定性中具有重要作用, 地上凋落物的输入显著促进了土壤团聚体的稳定性.本次采样结果中, 地上凋落物处理和氮添加并未显著影响土壤微生物生物量碳、氮及其比值, 由于微生物响应的敏感性和适应性, 本研究强调了未来关于微生物响应气候变化时空动态特征研究的重要性.
致谢: 感谢中国科学院植物研究所苏华老师、刘上石博士、冯银平硕士和北京林业大学周志勇老师在样品测试与分析中给予的帮助.
Allison S D, Vitousek P M. 2005. Responses of extracellular enzymes to simple and complex nutrient inputs[J]. Soil Biology and Biochemistry, 37: 937-944. DOI:10.1016/j.soilbio.2004.09.014 |
Belay-Tedla A, Zhou X, Su B, et al. 2009. Labile, recalcitrant, and microbial carbon and nitrogen pools of a tallgrass prairie soil in the US Great Plains subjected to experimental warming and clipping[J]. Soil Biology & Biochemistry, 41: 110-116. |
Berti A, Morari F, Dal Ferro N, et al. 2016. Organic input quality is more important than its quantity:C turnover coefficients in different cropping systems[J]. European Journal of Agronomy, 77: 138-145. DOI:10.1016/j.eja.2016.03.005 |
Blair G, Lefroy R, Lisle L. 1995. Soil carbon fractions based on their degree of oxidation, and the development of a carbon management index for agricultural systems[J]. Australian Journal of Agricultural Research, 46: 1459-1466. DOI:10.1071/AR9951459 |
Bucka F B, Kölbl A, Uteau D, et al. 2019. Organic matter input determines structure development and aggregate formation in artificial soils[J]. Geoderma, 354: 113881. DOI:10.1016/j.geoderma.2019.113881 |
Cotrufo M F, Wallenstein M D, Boot C M, et al. 2013. The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization:do labile plant inputs form stable soil organic matter?[J]. Global Change Biology, 19: 988-995. DOI:10.1111/gcb.12113 |
Davidson E A, Janssens I A. 2006. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change[J]. Nature, 440: 165-173. DOI:10.1038/nature04514 |
Dieckow J, Mielniczuk J, Knicker H, et al. 2005. Composition of organic matter in a subtropical Acrisol as influenced by land use, cropping and N fertilization, assessed by CPMAS 13C NMR spectroscopy[J]. European Journal of Soil Science, 56: 705-715. |
Dungait J A, Hopkins D W, Gregory A S, et al. 2012. Soil organic matter turnover is governed by accessibility not recalcitrance[J]. Global Change Biology, 18: 1781-1796. DOI:10.1111/j.1365-2486.2012.02665.x |
Forstner S J, Wechselberger V, Müller S, et al. 2019. Vertical redistribution of soil organic carbon pools after twenty years of nitrogen addition in two temperate coniferous forests[J]. Ecosystems, 22: 379-400. DOI:10.1007/s10021-018-0275-8 |
Frey S D, Knorr M, Parrent J L, et al. 2004. Chronic nitrogen enrichment affects the structure and function of the soil microbial community in temperate hardwood and pine forests[J]. Forest Ecology and Management, 196: 159-171. DOI:10.1016/j.foreco.2004.03.018 |
Frey S D, Nadelhoffer K J, LeMoine J M, et al. 2018. Molecular-level changes in soil organic matter composition after 10 years of litter, root and nitrogen manipulation in a temperate forest[J]. Biogeochemistry, 141: 183-197. DOI:10.1007/s10533-018-0512-4 |
郭魏, 齐学斌, 李平, 等. 2017. 不同施氮水平下再生水灌溉对土壤细菌群落结构影响研究[J]. 环境科学学报, 37(1): 280-287. |
Jaffrain J, Gérard F, Meyer M, et al. 2007. Assessing the quality of dissolved organic matter in forest soils using ultraviolet absorption spectrophotometry[J]. Soil Science Society of America Journal, 71: 1851-1858. DOI:10.2136/sssaj2006.0202 |
Janssens I A, Dieleman W, Luyssaert S, et al. 2010. Reduction of forest soil respiration in response to nitrogen deposition[J]. Nature Geoscience, 3: 315-322. DOI:10.1038/ngeo844 |
Jia Y, Yu G, He N, et al. 2014. Spatial and decadal variations in inorganic nitrogen wet deposition in China induced by human activity[J]. Scientific reports, 4: 3763. |
Lajtha K, Bowden R D, Crow S, et al. 2018. The detrital input and removal treatment (DIRT) network:Insights into soil carbon stabilization[J]. Science of the Total Environment, 640-641: 1112-1120. DOI:10.1016/j.scitotenv.2018.05.388 |
Lal R. 2004. Soil carbon sequestration to mitigate climate change[J]. Geoderma, 123: 1-22. DOI:10.1016/j.geoderma.2004.01.032 |
Le B Y, Prieto I, Roumet C, et al. 2018. Soil aggregate stability in Mediterranean and tropical agro-ecosystems:effect of plant roots and soil characteristics[J]. Plant and Soil, 424: 303-317. DOI:10.1007/s11104-017-3423-6 |
鲁如坤. 2000. 土壤农业化学分析方法[M]. 北京: 中国农业科学技术出版社.
|
Mambelli S, Bird J A, Gleixner G, et al. 2011. Relative contribution of foliar and fine root pine litter to the molecular composition of soil organic matter after in situ degradation[J]. Organic Geochemistry, 42: 1099-1108. |
Näthe K, Levia D F, Steffens M, et al. 2017. Solid-state 13C NMR characterization of surface fire effects on the composition of organic matter in both soil and soil solution from a coniferous forest[J]. Geoderma, 305: 394-406. DOI:10.1016/j.geoderma.2017.06.030 |
Norby R J, Warren J M, Iversen C M, et al. 2010. CO2 enhancement of forest productivity constrained by limited nitrogen availability[J]. Proceedings of the National Academy of Sciences of the United States of America, 107: 19368-19373. DOI:10.1073/pnas.1006463107 |
Pan Y, Birdsey R A, Fang J, et al. 2011. A large and persistent carbon sink in the world's forests[J]. Science, 333: 988-993. DOI:10.1126/science.1201609 |
Schippers P, Lurling M, Scheffer M. 2004. Increase of atmospheric CO2 promotes phytoplankton productivity[J]. Ecology Letters, 7: 446-451. DOI:10.1111/j.1461-0248.2004.00597.x |
Silva-Sánchez A, Soares M, Rousk J. 2019. Testing the dependence of microbial growth and carbon use efficiency on nitrogen availability, pH, and organic matter quality[J]. Soil Biology and Biochemistry, 134: 25-35. DOI:10.1016/j.soilbio.2019.03.008 |
Six J, Paustian K, Elliott E, et al. 2000. Soil structure and organic matter I. Distribution of aggregate-size classes and aggregate-associated carbon[J]. Soil Science Society of America Journal, 64: 681-689. DOI:10.2136/sssaj2000.642681x |
Sokol N W, Kuebbing S E, Karlsen-Ayala E, et al. 2019. Evidence for the primacy of living root inputs, not root or shoot litter, in forming soil organic carbon[J]. New Phytol, 221: 233-246. DOI:10.1111/nph.15361 |
Sun X, Tang Z, Ryan M G, et al. 2019. Changes in soil organic carbon contents and fractionations of forests along a climatic gradient in China[J]. Forest Ecosystems, 6: 1. DOI:10.1186/s40663-019-0161-7 |
Sun X L, Zhao J, You Y M, et al. 2016. Soil microbial responses to forest floor litter manipulation and nitrogen addition in a mixed-wood forest of northern China[J]. Scientific Reports, 6: 19536. DOI:10.1038/srep19536 |
Tian H, Lu C, Yang J, et al. 2015. Global patterns and controls of soil organic carbon dynamics as simulated by multiple terrestrial biosphere models:Current status and future directions[J]. Global Biogeochemical Cycles, 29: 775-792. DOI:10.1002/2014GB005021 |
Tian Q, Yang X, Wang X, et al. 2016. Microbial community mediated response of organic carbon mineralization to labile carbon and nitrogen addition in topsoil and subsoil[J]. Biogeochemistry, 128: 125-139. DOI:10.1007/s10533-016-0198-4 |
涂玉, 尤业明, 孙建新. 2012. 油松-辽东栎混交林地表凋落物与氮添加对土壤微生物生物量碳、氮及其活性的影响[J]. 应用生态学报, 23: 2325-2331. |
Vogel C, Babin D, Pronk G J, et al. 2014. Establishment of macro-aggregates and organic matter turnover by microbial communities in long-term incubated artificial soils[J]. Soil Biology & Biochemistry, 79: 57-67. |
von Lutzow M, Kogel-Knabner I, Ekschmitt K, et al. 2006. Stabilization of organic matter in temperate soils:mechanisms and their relevance under different soil conditions-a review[J]. European Journal of Soil Science, 57: 426-445. DOI:10.1111/j.1365-2389.2006.00809.x |
Wallenstein M D, Hall E K. 2012. A trait-based framework for predicting when and where microbial adaptation to climate change will affect ecosystem functioning[J]. Biogeochemistry, 109: 35-47. DOI:10.1007/s10533-011-9641-8 |
Wang R, Goll D, Balkanski Y, et al. 2017. Global forest carbon uptake due to nitrogen and phosphorus deposition from 1850 to 2100[J]. Global Change Biology, 23: 4854-4872. DOI:10.1111/gcb.13766 |
夏品华, 喻理飞, 寇永珍, 等. 2017. 贵州高原草海湿地土壤有机碳分布特征及其与酶活性的关系[J]. 环境科学学报, 37(4): 1479-1485. |
Xu S, Liu L L, Sayer E J, et al. 2013. Variability of above-ground litter inputs alters soil physicochemical and biological processes:a meta-analysis of litterfall-manipulation experiments[J]. Biogeosciences, 10: 7423-7433. DOI:10.5194/bg-10-7423-2013 |
Yang Y, Luo Y, Lu M, et al. 2011. Terrestrial C:N stoichiometry in response to elevated CO2 and N addition:a synthesis of two meta-analyses[J]. Plant and Soil, 343: 393-400. DOI:10.1007/s11104-011-0736-8 |
朱仁欢, 郑子成, 李廷轩, 等. 2018. 退耕植茶对川西低山丘陵区土壤活性有机碳组分的影响[J]. 环境科学学报, 38(2): 744-751. |