 2021, Vol. 41
2021, Vol. 41



2. 北京市水科学技术研究院, 北京 100048;
3. 中国科学院动物研究所, 北京 100101
2. Beijing Water Science and Technology Institute, Beijing 100048;
3. Institute of Zoology, Chinese Academy of Sciences, Beijing 100101
鱼类是水生生态系统的主要组成部分, 在维持区域生态系统平衡、保护水资源和水环境安全方面具有重要的指示作用(张春光等, 2011).鱼类群落结构变化是对其水域环境和人为影响变化的响应, 其相关过程的演变可能导致渔业功能退化, 也会使河流生态系统失去调控的重要功能, 因此, 保持河流鱼类群落结构的合理性是河流管理和生态系统恢复的关键(Pan et al., 2016).城市的快速发展对河流生态系统造成了严重的威胁, 有效协调城市发展与鱼类生物多样性保护之间的矛盾已成为重要前提(Paukert et al., 2011).
在水生态区域划分的研究中, 常把鱼类群落作为较大尺度水生态分区的指示生物, 其空间分布能够体现较大尺度上河流生境的梯度变化(Abell et al., 2008;Gao et al., 2015).例如, 王云涛等(2016)对影响太子河流域不同水生态区鱼类群落分布与河流健康的环境因子进行筛选, 结果表明, 水生态不同区域(上游山地区、中游丘陵区、下游平原区)处于不同的生态健康等级, 制订流域鱼类保护方案应当充分考虑不同水生态区特定环境因素的影响;王晓宁等(2018)通过空间聚类的方法, 将具有相似性鱼类群落特征的区域进行空间划分, 发现山区与平原不同区域河流的多样性指数与指标影响因子具有显著的差异性;何思璇等(2019)通过对防城河口湾海域进行渔业资源调查, 研究了河口湾鱼类群落结构及其空间和季节变化, 结果表明, 该海域鱼类群落存在一定程度的时空异质性, 影响鱼类群落结构的主要环境因子为水温、盐度和硝酸盐.不同的鱼类群落结构是对不同环境下地理特征的响应, 研究城市河流不同水生态区下的鱼类群落结构及环境影响因子, 了解生物群落在不同环境干扰压力下的退化机制, 有助于恢复群落多样性, 对后期河流生态修复具有重要意义(孟伟等, 2007;李艳利等, 2014;丁越岿等, 2016).
北运河是北京市重要的排水通道和连接京津冀地区的生态走廊, 在首都社会经济发展及生态系统的维系中具有极其重要的地位, 但经济的快速发展及人口数量的激增导致河道污水排量加大、水域生态空间压缩、水体自我修复能力弱, 最终使河流生态系统逐渐失去平衡(刘宇同等, 2019).前期对北运河的相关生态研究均是在自然状态下受到气象、地质与地势条件差异的影响, 生物在不同空间区域的群落组成、优势种表现出不同的特征(杨柏贺等, 2019;贺玉晓等, 2020);而基于人为活动因素干扰剧烈的非自然状态下, 对北运河鱼类资源调查及群落结构的研究较少, 特别是北运河上游南沙河段与下游城市副中心段(北关闸-甘棠闸), 代表了北运河近山区和平原不同水生态空间下的典型河段, 属于人工开挖疏浚河道且两岸为人工护坡, 具有较好的研究意义.因此, 本研究针对人为活动因素干扰剧烈的非自然状态下, 对比北运河上、下游不同河段鱼类群落组成及多样性特征的空间差异, 解析群落分布与环境因子的响应关系, 明确不同河段后期的生态修复方向, 为北运河流域污染治理及生态修复提供指导.
2 材料与方法(Materials and methods) 2.1 研究区概况北运河属于海河水系北部, 起源于北京燕山以南, 自西北向东南流经北京巿、河北省和天津市后入海河, 上游为山区丘陵地带, 中、下游为华北冲积平原.北京市境内的北运河河道长89.4 km, 流域面积为4293 km2, 流域内有大小支流110条, 年度出境水量为7亿m3.流域内属于温带大陆性季风气候, 多年平均降水量为611 mm, 降水量集中在汛期的6—9月.
南沙河发源于北京市昌平区西南部山区和海淀区西北部, 属于北运河水系上游, 主河道长22.21 km, 流域面积为220 km2, 河道为土渠梯形断面, 河深约为4.0~5.0 m, 多年平均降水量为594.8 mm, 承担海淀北部地区的排洪、灌溉、纳污等功能;近年来随着截污治污工作加强, 南沙河水质有所好转, 但水生态环境仍未达到相关功能要求.北运河城市副中心段属于北运河平原干流下游, 长11.4 km, 水势受季节影响显著, 河面宽阔河岸顺直, 河流城市化特征显著, 随着北京市城市副中心的确立与发展, 其承担的生态、景观、防洪等重要性功能日益凸显, 未来将逐渐成为流域内可持续发展的焦点(于洋等, 2012;崔驰飞等, 2019).
2.2 样点选取及样品采集于2019年10月上旬和下旬, 分别根据北运河上游南沙河段和下游城市副中心段调查水体和环境的具体情况选择调查地点, 其中, 南沙河段布设样点8个, 副中心段布设样点4个, 分别标记NS1、NS2、NS3、NS4、NS5、NS6、NS7、NS8和CS1、CS2、CS3、CS4, 其中, NS2、NS7、CS2点位未采集到数据(图 1、表 1).
 |
| 图 1 北运河上游南沙河段与下游城市副中心段采样点示意图 Fig. 1 Map of sampling points between upstream and downstream of the Beiyun River |
| 表 1 北运河上游南沙河段与下游城市副中心段采样点位置信息表 Table 1 Location information of sampling points between upstream and downstream of the Beiyun River |
选择15 m长的地笼作为采样工具, 采样时均自下游向上游行进.待样品采集后, 现场进行鉴定, 记录采集地点、时间、记录人等信息, 测量样品长度、体重、尾数, 并拍照.现场不能鉴定的疑难种, 用福尔马林固定保存, 带回实验室进一步鉴定.
2.3 鱼类标本鉴定标本鉴定与分类主要依据《北京及其邻近地区的鱼类》(张春光等, 2013)、《中国内陆鱼类物种与分布》、《中国鱼类系统检索》.
2.4 数据处理和分析 2.4.1 优势种的确定将渔获物中单个物种个体数占渔获物总数10%以上的物种定为优势种(郁尧山等, 1986).
2.4.2 群落多样性分析Shannon-Wiener多样性指数(H)、Margalef丰富度指数(D)和Pielous均匀性指数(J)的计算公式如下:
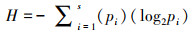
|
(1) |
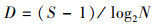
|
(2) |
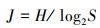
|
(3) |
式中, S为采集所得鱼类标本的种类总数;N为采集所得鱼类标本总个体数;pi为第i种鱼类占总数的比例(刘斌等, 2013).
2.4.3 数据分析应用SPSS 17.0软件对北运河上游南沙河段与下游城市副中心段鱼类群落多样性、丰富度、均匀度指数做箱图对比;应用SPSS 17.0软件中Pearson相关性分析分别检验鱼类群落各指数与环境因子的相关性;应用Canoco 4.5软件对鱼类群落数据和环境因子数据进行典范对应分析(Canonical Correspondence Analysis CCA), 所有数据都进行lg(x+1)转换, 再进行9999次蒙特卡洛循环检验(Monte Carlo Test, MCT), 并将单个因子贡献率的p值小于0.05的环境因子作为显著影响鱼类群落分布的水环境因子(王晓宁等, 2018).
3 结果(Results) 3.1 北运河南沙河段与副中心段鱼类群落组成北运河上游南沙河段与下游城市副中心段鱼类群落组成见表 2.南沙河段与副中心段共调查鱼类14种, 属于3目5科11属, 鲤形目(Cypriniformes)种类最多, 有9种, 占总物种数的64.2%;其次是鲈形目(Perciformes), 有4种, 占28.57%;合鳃鱼目(Synbranchiformes)有1种, 占7%.从科级水平上看, 鲤科种类最多有7种, 占总物种数的50%;鳅科(Cobitidae)、虾虎鱼科(Gobiidae)、塘鳢科(Eleotridae)和合鳃鱼科(Synbranchidae)种类较少.分河段来看, 南沙河段鱼类共调查到7种, 以占该河段92%的麦穗鱼为优势种, 副中心段调查到鱼类种数13种, 大鳞副泥鳅、麦穗鱼、泥鳅分别占该河段的40%、24%、20%, 均为优势种.
| 表 2 北运河上游南沙河段与下游城市副中心段鱼类群落组成 Table 2 Fish community composition between upstream and downstream of the Beiyun River |
北运河上游南沙河段与下游城市副中心段鱼类群落多样性指数变动幅度不同, 南沙河段多样性指数、丰富度指数、均匀度指数分别处于0.04~0.75、0.13~0.62、0.04~0.37之间, 副中心段多样性指数、丰富度指数、均匀度指数分别处于1.83~2.54、0.9~1.49、0.61~0.73之间, 表现为南沙河段多样性指数、均匀度指数变动幅度较大, 副中心段丰富度指数变动幅度较大的特征.南沙河段与副中心段各指数的对比箱图见图 2, 表现出上游南沙河段鱼类平均Shannon-Wiener多样性指数(0.33)、Margalef丰富度指数(0.31)、Pielou均匀度指数(0.17)均低于下游城市副中心段(2.11、0.98、0.93)的特征.
 |
| 图 2 北运河上游南沙河段(NS)与下游城市副中心段(CS)Shannon-Wiener指数、Margalef指数、Pielou指数箱图对比 Fig. 2 Comparison of Shannon Wiener index, Margalef index and Pielou index between upstream and downstream of the Beiyun River |
CCA环境因子分析结果见图 3, 北运河上游南沙河段与下游城市副中心段鱼类群落均受到ρ(TN)、ρ(TP)、ρ(NH4+-N)、ρ(COD)、ρ(NO3--N)环境因子的影响, 不同河流影响鱼类群落分布的环境因子有所差异:上游南沙河段中ρ(NO3--N)、ρ(COD)和ρ(TP)是显著影响鱼类群落分布的环境因子, 优势种麦穗鱼(Pseudorasbora parva)与ρ(NO3--N)、子陵吻虾虎鱼(Rhinogobius giurinus)与ρ(COD)和ρ(TP)分别呈显著正相关;下游城市副中心段ρ(NH4+-N)、ρ(TP)是显著影响鱼类群落分布的环境因子, 鱼类群落分布表现出棒花鱼(Abbottina rivularis)与ρ(NH4+-N)、黄鳝(Monopterus albus)、鲫(Carassius auratus)与ρ(TP)分别呈显著正相关.
 |
| 图 3 北运河上游南沙河段与下游城市副中心段鱼类群落分布与环境因子的典范对应分析结果 (图中TN表示ρ(TN), 其余TP、NH4+-N、COD、NO3--N类同) Fig. 3 CCA of species-environmental relationships between upstream and downstream of the Beiyun River |
Shannon-Wiener多样性指数、Margalef丰富度指数、Pielou均匀度指数与环境因子的Pearson相关性分析见表 3.结果表明, 各指数在P < 0.01水平上与ρ(NO3--N)呈显著负相关, 在P < 0.05水平上与ρ(TP)、ρ(TN)呈显著负相关, 表明各指数受到水质物理化学指标影响强烈.
| 表 3 多样性指数、丰富度指数、均匀度指数与环境因子的Pearson相关性分析 Table 3 Pearson correlation analysis of diversity index, richness index, evenness index and environmental factors |
杜龙飞等(2019)、张春光等(2011)分别对北京市各大水系主要河流的鱼类群落空间格局特征进行了研究, 李雪健等(2018)、杨文波等(2008)、王所安等(1987)分别对北京市拒马河、潮白河、永定河的鱼类资源进行了调查, 与历史资料相比, 受河流缺水、沙化、污染等生态条件退化影响, 整体反映出北京市各大水系鱼类资源呈退化的趋势.本研究中北运河上游南沙河段与下游城市副中心段共调查到鱼类14种, 基本是麦穗鱼、泥鳅等具有高度忍耐力的种类, 属于杂食性和肉食性鱼类, 没有植食性鱼类, 表现出高抗污染、广布型的特征;区域内鱼类以鲤形目为主并占绝对优势, 鲤形目中以鲤科为主要类群且占到总种数的一半以上, 与北京及我国北方整体鱼类群落组成特点相吻合.北运河上下游山区与平原不同的区域环境反映为鱼类指示种的生态特征差异:北运河上游近山区南沙河段调查到鱼类7种, 属于2目3科5属, 与山区河流拒马河相比在鱼类种属数量上较少, 均表现出以麦穗鱼为标志性优势物种的特点;麦穗鱼属于低营养级的中低层小型鱼类, 生长速度快、生命周期短, 常生活于较缓静浅水区;北京地区山区河流中常见的优势种包括鲤科的拉氏鱥、马口鱼、宽鳍鱲, 鳅科的高原鳅属、北方须鳅、花斑副沙鳅在此次调查中均未发现, 典型反映了南沙河段鱼类群落优势种单一和小型的特征.北运河下游城市副中心段调查到鱼类13种, 属于3目5科11属, 同与其临近的潮白河水系以宽鳍、高体鳑鲏和拉氏鱥为优势种相比, 副中心段以大鳞副泥鳅和泥鳅为指示性优势种且种类数量偏少;大鳞副泥鳅主要来自于人工引入或经水工设施扩散分布的结果, 泥鳅种多广泛分布于平原地区水体中, 对环境变化及水温的耐受力较强, 常被视为耐污性鱼类;外来物种河川沙塘鳢属于长江流域物种, 主要分布于我国长江中下游、钱塘江水系和闽江水系, 黄鳝在淮河以北数量较少, 常营穴居生活在池塘、河渠等淤泥质水底层, 该类物种进入栖息地后与原有物种争夺食物和生存空间, 可能对河段鱼类群落造成一定干扰.
4.2 北运河上下游不同河段鱼类群落多样性空间特征分析生物空间分布格局是群落在异质性生境和人类活动干扰长期影响过程中, 逐渐形成的一种适应性特征(帅方敏等, 2017).在无人类活动干扰的情况下, 河流下游水面变宽、水量变大, 为生物提供的生境类型更加丰富, 鱼类多样性增多(王忠锁等, 2006).鱼类群落的丰富度和多样性指数主要受其生境的异质性和稳定性影响, 均匀度指数可用来衡量群落中每种鱼生物量分类的均匀程度, 在一定程度上反映鱼类生物群落的稳定(Tammi et al., 1999;杨富亿等, 2012).Magurran(1988)提出多样性指数的一般范围是1.5~3.5, 就北京地区各大水系整体而言, 北运河水系中鱼类群落多样性指数、均匀度指数均低于潮白河水系和永定河水系(李雪健等, 2018).本研究中北运河上游南沙河段Shannon-Wiener多样性指数、Margalef丰富度指数、Pielou均匀度指数均低于下游城市副中心段, 主要与鱼类群落单一小型、栖息地特征及人类活动等因素有关:鱼类种类数量较少、优势种的单一小型导致生物多样性指数变幅大且数值较低, 这与毛志刚等(2011)的研究结果相一致.北运河上游南沙河起源于山区丘陵地带, 属于典型北方河道, 河流河道窄、河床浅, 影响河流的异质性和稳定性;下游城市副中心段地势平坦且水域面积宽阔, 属于北运河干流, 河流较长、支流较多, 日常径流量为300万m3·d-1, 城市段渠化现象较为普遍, 生境类型单一, 植被系统受到严重破坏且覆盖率不高, 影响生物生存的栖息地环境.除此之外, 近年来北京市快速城市化过程导致河流生态环境质量日渐下降, 水生生物多样性受到严重威胁(邢迎春等, 2007);特别是北运河流域内一系列水利工程、闸坝调控措施对河流生境造成一定的扰动, 暂未形成稳定的河流生态系统;沿河的土地利用、河道治理对鱼类群落有严重影响和危害, 进而导致群落多样性较低, 反映出城市化显著影响着北运河鱼类群落的空间多样性特征.
4.3 北运河上下游不同河段鱼类群落分布与环境因子的响应关系分析环境因素在很大程度上影响着鱼类的群落丰度, 如水体中污染物、悬浮颗粒物含量及富营养化改变等都是至关重要的因子(Paramo et al., 2003;Van et al., 2004), 水质指标对鱼类群落结构和分布的影响主要是通过改变鱼类物种的生理指标进而影响整个鱼类群落(王云涛等, 2016).相关研究表明(张勇等, 2012;李捷等, 2012), ρ(TN)、ρ(TP)等环境因子对河流鱼类群落和底栖动物分布都具有较大的影响.本研究中北运河南沙河段中影响鱼类群落分布的环境因子较副中心段更多, 表明该区域鱼类群落对水质环境因子更为敏感.由于南沙河支流来水水质较差, 黑臭淤积底泥释放的有机物污染、每年汛期形成的溢流污染, 导致南沙河长期经受污水影响(刘爱华, 2017), 特别是干流下游水质超标直接影响到鱼类群落分布.截至2019年7月, 温榆河、北运河干流仍存在部分排水口水质不达标的现象, 每年汛期有超过30%的河段会产生较为严重的水华, 在实地调查中副中心段河道底质多为黑臭淤泥, 重点断面水质状况主要体现为NH4+-N指标超标, NH4+-N浓度的变化规律体现了人类活动对河流水质的影响, 也间接反映鱼类群落空间分布与人类活动密切相关.同样, 环境因子的改变也是引起鱼类多样性变化的原因(张涛等, 2009), 营养盐浓度高的水域浮游生物资源丰富, 水体中藻类悬浮颗粒物较多, 透明度低, 会抑制鱼类非耐污种的生长, 有利于小型浮游生物食性的鱼类迅速形成优势群落, 从而降低该水域的鱼类生物多样性(Brown et al., 2000).本研究相关性分析结果表明, 营养盐类等有害物质的增多将直接导致水体中鱼类群落各类指数的降低, 这与陈辉等(2013)研究城市内河中鱼类多样性对环境因子的响应结果基本一致, 表明北运河上下游河段不同的水质状况与鱼类群落分布和多样性特征有一定关联性.
4.4 北运河鱼类群落栖息地恢复及资源保护建议北运河上游南沙河段与下游城市副中心段均属人为扰动比较剧烈的区域, 针对两段河道水质差、底泥黑臭现象严重的特点, 南沙河段后期的鱼类栖息地生态恢复重点应该以水质改善为主, 在适宜地区恢复自然生境或人工营造适宜鱼类繁衍的小环境, 增加环境异质性逐渐恢复鱼类群落的多样性;副中心段后期鱼类栖息地生态恢复应该在水质改善基础上, 集中于对鱼类多样性的保护和丰富生境的营造.进一步加强水环境的治理将对鱼类群落及生物多样性保护起到重要的作用, 管理者应根据不同河段的生态特性, 对鱼类采取有针对性的保护措施:在水质恢复方面, 北运河上游南沙河段应考虑进一步加强截污治污, 并结合水土保持、植物对污染物的削减等措施建设植被缓冲带进行面源污染物防控, 加强溢流污染治理, 在入河排水口等区域通过建设河道型湿地、增加水生植被等生态措施净化水体;下游城市副中心段水质应重点通过开展截污治污工程实现对排水口的治理, 同时加强生态湿地建设, 进一步构建复杂生境系统, 提升水体净化能力和生态系统结构稳定性.在水量调配方面, 优先采用再生水为水源, 制定科学生态补水方案, 选择适宜的时段、环境、物种进行增殖放流等生物管控措施, 同时定期对河道进行生态清淤等综合治理.在生态监测方面, 逐年开展对受水区鱼类多样性变化的跟踪调查, 动态监测鱼类多样性、群落结构等的长期变化, 密切关注外来鱼类, 严防物种入侵对原有群落生存空间的干扰.在政策制度方面, 加强野生鱼类资源保护的宣传教育;尽快建立野生鱼类种质资源库, 为今后开展资源恢复工作提供后备原生鱼种;根据实际情况, 建立区域性禁渔期制度, 在此期间流域内严禁一切渔业生产活动, 保证野生鱼类有自然生产补充资源的时机.
5 结论(Conclusions)1) 北运河调查到鱼类分属3目5科11属14种, 表现出种属偏低、高抗污染、广布型的特征:其中, 上游南沙河段鱼类共调查到7种, 以麦穗鱼为优势种, 反映鱼类群落单一和小型的特征, 下游城市副中心段调查到鱼类种数13种, 以大鳞副泥鳅、麦穗鱼、泥鳅为优势种, 河川沙塘鳢为外来物种.
2) 北运河鱼类群落多样性指数整体偏低, 上游南沙河段Shannon-Wiener多样性指数、Margalef丰富度指数、Pielou均匀度指数均低于下游城市副中心段鱼类群落各指数, 主要与鱼类群落单一小型、栖息地环境及人类活动等因素有关.
3) 影响鱼类群落分布的环境因子中, ρ(NO3--N)、ρ(TP)、ρ(COD)对上游南沙河段鱼类群落的分布影响较大, ρ(NH4+-N)、ρ(TP)对下游城市副中心段鱼类群落分布影响较大, 多样性、丰富度和均匀度指数均表现出与ρ(TP)、ρ(TN)、ρ(NO3--N)呈显著负相关, 表明上下游河段不同的水质状况与鱼类群落分布和多样性特征有一定关联性.
4) 北运河上游南沙河段与下游城市副中心段受人类活动影响显著, 主要表现在河道水质差、底泥黑臭现象严重方面, 南沙河段后期的鱼类栖息地生态恢复重点应该以水质改善为主, 副中心段后期鱼类栖息地生态恢复应该在水质改善的基础上集中于对鱼类多样性的保护和丰富生境的营造, 生态修复措施围绕水质改善、生态补水、群落监测、政策管控方面进行.
Abell R, Tihemem L, Revenga C, et al. 2008. Freshwater ecoregions of the world:a new map of biogeographic units for freshwater biodiversity conservation[J]. Bioscience, 58(5): 403-414. DOI:10.1641/B580507 |
Brown L R. 2000. Fish communities and their associations with environmental variables, lower San Joaquin River Drainage, California[J]. Environmental Biology of Fishes, 57(3): 251-269. DOI:10.1023/A:1007660914155 |
陈辉, 王继华, 刘翔, 等. 2013. 常州市城市内河常见鱼类资源及保护的研究[J]. 中国农学通报, 29(32): 66-70. |
崔驰飞, 祁诗月, 郝明, 等. 2019. 城市黑臭水体治理技术研究-以北京市海淀区南沙河流域为例[J]. 北京水务, (2): 12-16. |
丁越岿, 张洪, 单保庆. 2016. 海河流域河流空间分布特征及演变趋势[J]. 环境科学学报, 36(1): 47-54. |
杜龙飞, 徐建新, 李彦彬, 等. 2019. 北京市主要河流鱼类群落的空间格局特征[J]. 环境科学研究, 32(3): 447-457. |
Gao X, Ding S, Zhang Y, et al. 2015. Exploring the relationship among land-use, riparian habitat quality, and biological integrity of a fish community[J]. Acta Ecologica Sinica, 35(21): 7198-7206. |
何思璇, 何斌源. 2019. 防城河口湾鱼类群落结构及其与环境因子关系研究[J]. 热带海洋学报, 38(5): 86-97. |
贺玉晓, 刘天慧, 任玉芬, 等. 2020. 北运河秋冬季浮游植物群落结构特征及影响因子分析[J]. 环境科学学报, 40(5): 1710-1721. |
李捷, 李新辉, 贾晓平, 等. 2012. 连江鱼类群落多样性及其与环境因子的关系[J]. 生态学报, 32(18): 5795-5805. |
李雪健, 郭久波, 牛诚祎, 等. 2018. 北京市潮白河流域鱼类物种组成的历史演变和多样性现状[J]. 动物学杂志, 53(3): 375-388. |
李艳利, 李艳粉, 徐宗学. 2014. 影响浑太河流域鱼类群落结构的不同尺度环境因子分析[J]. 环境科学, 35(9): 3504-3512. |
刘爱华. 2017. 北京市南沙河流域水环境质量评价研究[J]. 中国环保产业, (6): 63-67. DOI:10.3969/j.issn.1006-5377.2017.06.018 |
刘斌, 张远, 渠晓东, 等. 2013. 辽河干流自然保护区鱼类群落结构及其多样性变化[J]. 淡水渔业, 43(3): 49-55. DOI:10.3969/j.issn.1000-6907.2013.03.009 |
刘宇同, 杨伟超, 杨丽娜, 等. 2019. 北运河流域水生态恢复与保护的实践探索[J]. 北京水务, (3): 57-62. |
Magurran A E.1988.Ecological diversity and its measurement[M].Springer, Dordrecht: 92(12): 2285-2298
|
毛志刚, 谷孝鸿, 曾庆飞, 等. 2011. 太湖鱼类群落结构及多样性[J]. 生态学杂志, 30(12): 2836-2842. |
孟伟, 张远, 郑丙辉. 2007. 辽河流域水生态分区研究[J]. 环境科学学报, 27(6): 911-918. DOI:10.3321/j.issn:0253-2468.2007.06.004 |
Paramo J, Quinones R A, Ramirez A, et al. 2003. Relationship between abundance of small pelagic fishes and environmental factors in the Colombian Caribbean Sean analysis based on hydroacoustic information[J]. Aquatic Living Resources, 16(3): 239-245. DOI:10.1016/S0990-7440(03)00043-3 |
Pan B Z, Yuan J P, Zhang X H, et al. 2016. A review of ecological restoration techniques in fluvial rivers[J]. International Journal of Sediment Research, 31: 110-119. DOI:10.1016/j.ijsrc.2016.03.001 |
Paukert C P, Pitts K L, Whittier J B, et al. 2011. Development and assessment of a landscape-scale ecological threat index for the Lower Colorado River Basin[J]. Ecological Indicators, 11(2): 304-310. DOI:10.1016/j.ecolind.2010.05.008 |
帅方敏, 李新辉, 刘乾甫, 等. 2017. 珠江水系鱼类群落多样性空间分布格局[J]. 生态学报, 37(9): 3182-3192. |
Tammi J, Lappalainen A, Mannio J, et al. 1999. Effects of eutrophication on fish and fisheries in Finnish lakes:a survey based on random sampling[J]. Fisheries Management and Ecology, 6(3): 173-186. DOI:10.1046/j.1365-2400.1999.00152.x |
王晓宁, 彭世贤, 张亚, 等. 2018. 滦河流域鱼类群落结构空间异质性与影响因子分析[J]. 环境科学研究, 31(2): 273-282. |
王云涛, 张远, 高欣, 等. 2016. 太子河流域不同水生态区鱼类群落分布与环境因子的关联性[J]. 环境科学研究, 29(2): 192-201. |
王所安, 柳殿钧, 曹玉萍. 1987. 永定河系的环境条件和自然鱼类资源[J]. 河北大学学报(自然科学版), (4): 36-41+98-99. |
王忠锁, 陈明华, 吕偲, 等. 2006. 鄱阳湖银鱼多样性及其时空格局[J]. 生态学报, 26(5): 1337-1344. |
Van Sickle, Baker J J, Herlihy A, et al. 2004. Projecting the biological condition of streams under alternative scenarios of human land use[J]. Ecological Applications, 14(1): 368-380. |
邢迎春, 赵亚辉, 李高岩, 等. 2007. 北京市怀沙-怀九河市级水生野生动物保护区鱼类物种多样性及其资源保护[J]. 动物学杂志, (1): 29-37. |
杨柏贺, 马思琦, 王汨, 等. 2019. 北运河水系大型底栖动物摄食功能群多样性及时空分布特征[J]. 河南师范大学学报(自然科学版), 47(1): 105-111. |
杨富亿, 吕宪国, 娄彦景, 等. 2012. 莫莫格国家级自然保护区的鱼类多样性[J]. 湿地科学, 10(2): 214-222. |
杨文波, 李继龙, 李绪兴, 等. 2008. 拒马河北京段鱼类组成及其多样性[J]. 上海水产大学学报, (2): 175-181. |
于洋.2012.北运河水体中氨氮的氧化过程及微生物响应特征[D].北京: 首都师范大学
|
郁尧山, 张庆生, 陈卫明, 等. 1986. 浙江北部岛礁周围海域鱼类优势种及其种间关系的初步研究[J]. 水产学报, 10(2): 137-149. |
张春光, 赵亚辉, 邢迎春, 等. 2011. 北京及其邻近地区野生鱼类物种多样性及其资源保育[J]. 生物多样性, 19(5): 597-604. |
张春光, 赵亚辉. 2013. 北京及其邻近地区的鱼类[M]. 北京: 科学出版社.
|
张涛, 庄平, 刘健, 等. 2009. 长江口崇明东滩鱼类群落组成和生物多样性[J]. 水生态学杂志, 28(10): 2056-2062. |
张勇, 刘朔孺, 于海燕, 等. 2012. 钱塘江中游流域不同空间尺度环境因子对底栖动物群落的影响[J]. 生态学报, 32(14): 4309-4317. |