 2021, Vol. 41
2021, Vol. 41



堆肥是微生物主导的矿化-腐殖化的耦合过程, 腐殖化过程将有机物质合成稳定的腐殖质(李波等, 2017), 而矿化过程将有机物转化成CH4、CO2、NH3和NOx等气体排入环境(黄华等, 2013), 从而导致碳氮损失.这不仅造成环境污染, 也会导致堆肥产品质量下降(Awasthi et al., 2016).因此, 如何减少堆肥过程中碳氮的损失对减少堆肥的二次环境污染, 提高堆肥质量具有重要意义.
堆肥过程受到C/N、湿度、温度、pH和添加剂等因素的深刻影响(Li et al., 2016).添加剂通过改善堆肥物料结构, 促进微生物的新陈代谢和有机质的降解, 成为堆肥过程的主要影响因素之一(Lei et al., 2000).堆肥添加剂包括有机类(生物质炭、木炭和竹炭等)和无机类(凹凸棒、沸石和过磷酸钙等).其中, 生物质炭是生物质在缺氧条件下通过热化学转化得到的固态产物(IBI, 2012), 具有多孔、强吸附力和高化学稳定性等特点(Gul et al., 2015;Luo et al., 2016), 可用以吸附氮素、减少有害气体排放而被广泛用作堆肥添加剂(贺琪等, 2005;Nelissen et al., 2012);凹凸棒具有成本低、安全和环保等优势, 以及强的吸附能力(李荣华, 2020), 可有效减少堆肥过程中氨气的挥发从而减少氮素的损失(Pan et al., 2019).但现有关于添加剂的研究主要集中在减少堆肥过程中养分损失的结果方面, 有关其微生物机理方面的研究相对较少.堆肥是微生物学过程, 添加剂如何驱动堆肥中优势微生物菌群的演替而影响碳氮转化过程, 从而减少养分损失的机理目前尚不明确.
因此, 本研究以鸡粪和玉米秸秆为原料, 不同含量的生物质炭和凹凸棒为添加剂进行堆肥, 探讨不同添加剂对堆肥中碳氮损失的作用, 同时利用16S rRNA基因测序分析不同添加剂对堆肥过程中优势微生物群落演替的影响, 通过优势微生物菌群与碳氮损失之间的相关性分析, 阐明添加剂影响堆肥过程中碳氮损失的微生物学机理, 从而探索堆肥减排增效的可能途径, 为减少堆肥过程中养分损失, 提高堆肥质量, 减少堆肥二次环境污染提供理论依据.
2 材料和方法(Materials and methods) 2.1 堆肥材料鸡粪和玉米秸秆原料均取自甘肃省会宁县钟岔村的农场, 玉米秸秆粉碎成1 cm左右;生物质炭(玉米秸秆为原料)购自北京三聚环保有限公司, 凹凸棒来源于甘肃白银.各材料基本性状见表 1.
| 表 1 原材料性质 Table 1 Properties of raw materials |
试验设置5个处理, 分别为CK(鸡粪+玉米秸秆)、BC1(鸡粪+玉米秸秆+5%(干重)生物质炭)、BC2(鸡粪+玉米秸秆+10%(干重)生物质炭)、PG1(鸡粪+玉米秸秆+5%(干重)凹凸棒))和PG2(鸡粪+玉米秸秆+10%(干重)凹凸棒).按照鸡粪和玉米秸秆的碳、氮含量和含水量, 称取鸡粪400 kg(鲜重), 并计算达到C/N为25∶1、含水量为60%±1%所需要的秸秆量(鲜重), 然后将计算好的秸秆量加入到称好的鸡粪中, 充分混匀;再按照不同添加剂的添加量加入添加剂, 调整含水量为60%±1%, 混匀装箱至箱体的100%(60 cm×60 cm×45 cm的泡沫箱), 敞口放置在校园带彩钢板顶的室外进行堆肥, 堆肥时间为2019年10月6—27日.每个处理重复3次, 所有堆肥箱每3 d翻堆一次, 翻堆前后均称重.分别于第1、4、7、10、13、16、19和22 d翻堆并采集样品, 采样前充分混匀, 采用多点取样混合取样500 g左右.每个样品分3份, 新鲜样品存储于4 ℃条件下进行发芽指数(GI)分析, 风干样品粉碎过0.1 mm筛用以全氮、全磷、全钾和总有机碳分析;真空包装的样品存储在-80 ℃冰箱中用于高通量测序, 根据碳氮损失量的测定数据, 选择碳氮损失更少的BC2、PG2和CK共3个处理的第1 d(初始期)、第4 d(高温期)、第10 d(降温期)、第22 d(腐熟期)样品送至北京百迈客生物技术有限公司进行基因测序.
2.3 理化指标测定每天9:00和15:00用温度计测量各堆心及环境温度, 记录并取平均值.含水率的测定采用烘干法, 全氮(TN)、全磷(TP)、全钾(TK)和有机质(OM)测定的具体方法参照中华人民共和国农业行业标准(NY525—2012)有机肥料行业标准.碳损失([C-loss])和氮损失([N-loss])按照下式计算(Zhang et al., 2020):
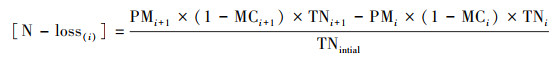
|
(1) |
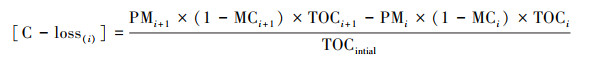
|
(2) |
式中, PM为堆体质量(kg), MC为含水量, TN为全氮含量(g·kg-1), TOC为总有机碳含量(g·kg-1), i=1~5代表采样次数.
堆肥样品与蒸馏水按质量比1∶10充分混匀, 摇床摇2 h后5000 r·min-1离心15 min, 吸取5 mL上清液加入到已铺有滤纸的无菌培养皿中;在滤纸上整齐摆放大小均匀的小白菜种子30粒, 同时以蒸馏水做空白对照, 放入25 ℃的培养箱中培养48 h后, 测定种子的发芽率和根长, 根长使用游标卡尺进行测量;根据公式计算发芽指数(GI)(Zucconi et al., 1981):
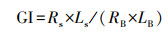
|
(3) |
式中, Rs为样品的发芽率, Ls为样品的总根长(cm), RB空白的发芽率, LB为空白的总根长(cm).
2.4 DNA提取和16S rRNA基因测序分析DNA采用MN NucleoSpin 96 Soi试剂盒按照说明提取, V3~V4区16S rRNA通用引物338F(5′- ACTCCTACGGGAGGCAGCA-3′)和806R (5′-GGACTACHVGGGTWTCTAAT-3′)进行PCR扩增.PCR反应条件:95 ℃预变性5 min;95 ℃变性30 s, 50 ℃退火30 s, 72 ℃延伸40 s, 25个循环;具体循环情况, 72 ℃, 7 min;4 ℃条件下保存.扩增产物进行纯化、定量和均一化形成测序文库, 建好的文库先进行文库质检, 质检合格的文库用Illumina HiSeq 2500进行双端测序, 得到原始图像数据文件, 经碱基识别分析转化为原始测序序列.细菌物种注释数据库选用Unite(http://unite.ut.ee/index.php).
2.5 数据分析用Excel 2019进行折线图绘制, 用百迈客生物云计算平台(BMKCloud)内置QIIME软件进行Beta多样性分析, 根据距离矩阵获得bray_curtis距离下的样品NMDS分析结果, 利用R语言Vegan包绘制相关性热图和进行冗余分析(RDA), 使用PICRUSt软件进行KEGG代谢途径的差异分析.
3 结果和分析(Results and analysis) 3.1 添加剂对堆肥腐熟度特性的影响温度是反映堆肥过程中微生物活性、有机物降解率和堆肥无害程度的重要指标(Gao et al., 2015;吴梦婷等, 2020).本研究5个处理的腐熟度相关指标见图 1.由图可知, 5个处理均在第4 d进入高温阶段(>50 ℃), 并持续7 d以上, 满足堆肥过程中的无害化温度要求(Bernal et al., 2009).PG1和PG2处理的堆肥温度在第10 d就低于60 ℃, 而CK、BC2和BC1处理的堆肥温度在第12 d才低于60 ℃, 添加凹凸棒导致C/N降低, 不能为微生物提供充足的养分, 使微生物活性降低导致温度快速下降(林小凤等, 2008).种子发芽指数(GI)是综合评价堆肥腐熟程度的指标之一(Wang et al., 2004).5个处理的GI值随堆肥进行逐渐上升(图 1b), 堆肥21 d时均达到了80%的堆肥腐熟标准(李丹阳等, 2020).由上面2个参数可知, 5个处理均达到了堆肥腐熟的标准.
 |
| 图 1 不同处理的温度(a)和发芽指数(b)变化趋势 Fig. 1 Trends in temperature (a) and GI (b) at different treatments |
随着堆肥的进行, 由于氨挥发和氮氧化物的产生造成氮素大量损失(黄华等, 2013), 5个处理的全氮含量都显著降低(图 2a), 当堆肥结束时, CK、BC1、BC2、PG1和PG2处理的全氮含量分别下降了18.08%、17.71%、13.61%、14.47%和15.43%.5个处理的全磷和全钾含量都显著增加(图 2b、2c), CK、BC1、BC2、PG1和PG2处理的全磷和全钾含量分别增加了36.08%、49.40%、50.00%、40.20%、42.70%和56.25%、66.89%、68.71%、58.39%、60.24%.5个处理的有机质含量均呈逐渐下降趋势(图 2d), CK、BC1、BC2、PG1和PG2处理的有机质含量分别下降了9.93%、7.86%、8.59%、9.40%和9.41%.到堆肥结束时, CK、BC1、BC2、PG1和PG2的氮、磷、钾总量分别为5.52%、5.2%、5.17%、5.43%和5.41%, 有机质含量分别为63.95%、65.85%、66.82%、64.89%和65.40%, 都高于我国有机肥质量标准(NY 525-2012).
 |
| 图 2 不同处理的全氮(a)、全磷(b)、全钾(c)和有机质(d)含量变化趋势 Fig. 2 Trends of total nitrogen(a), total phosphorus(b), total potassium(c) and organic matter(d) under different treatments |
碳氮损失会严重影响堆肥质量(Hong et al., 2014.5个处理的碳氮累积损失表现出相同趋势(图 3), 前3 d为快速损失阶段, 3~14 d为相对缓慢阶段, 14~22 d为稳定阶段.堆肥结束时, CK、BC1、BC2、PG1和PG2处理的累积碳损失分别为18.25%、16.68%、16.05%、17.88%和17.13%, 氮损失分别为23.24%、19.86%、18.56%、21.29%和20.63%.相对于CK, BC1、BC2、PG1、PG2处理的碳和氮损失分别减少了8.60%、12.05%、2.03%、6.14%和14.54%、20.14%、8.40%、11.23%.PG2和BC2处理的碳氮损失都低于BC1和PG1处理, 说明添加10%的生物质炭和凹凸棒更有利于减少碳氮损失, 而添加10%生物质炭的效果最佳.
 |
| 图 3 不同种处理的累积氮(a)和累积碳(b)损失 Fig. 3 Cumulative nitrogen loss(a) and cumulative carbon loss(b) under different treatments |
NMDS分析是一种将多维空间的研究对象简化到低维空间进行定位、分析和归类, 同时又保留对象间原始关系的数据分析方法(殷亚楠, 2018).由图 4可知, 在堆肥过程中, CK、BC2和PG2处理的细菌群落随堆肥进程明显地分割开来, 表明堆肥温度会显著影响细菌群落结构;在堆肥同一时期, CK、BC2和PG2处理间明显分割开来, 且随堆肥进程差异越明显, 表明添加剂显著影响了堆肥的细菌群落结构.从堆肥初始到结束, 除降温期(C)BC2与CK更接近外, 其余时期BC2均比PG2更远离CK, 表明生物质炭的添加对堆肥过程中细菌群落结构的影响远大于凹凸棒, 这与图 5的结果相一致.
 |
| 图 4 不同阶段3个处理组样品16S rRNA测序数据的NMDS分析 (CK、BC和PG分别代表CK处理、BC2处理和PG2处理, A、B、C、D分别代表堆肥的初始期、高温期、降温期和腐熟期, 1、2、3代表每个处理的3次重复) Fig. 4 NMDS analysis of 16S RNA sequencing data of three treatment groups at different stages (CK, BC and PG represent CK treatment, BC2 treatment and PG2 treatment, respectively.A, B, C, and D represent the initial period, high temperature period, cooling period and maturity period of compost, 1, 2, and 3 represent 3 repetitions of each treatment) |
 |
| 图 5 3种处理的细菌群落在属水平上的演替 (各处理编号后的A、B、C、D分别代表堆肥的初始期、高温期、降温期和腐熟期) Fig. 5 Succession of bacterial communities at genus level for the three treatments |
属水平上的细菌种类及相对丰度更能反映堆肥中微生物群落结构的变化(张嘉超, 2009), CK、BC2和PG2处理在4个堆肥阶段属水平上的微生物群演替如图 5所示.
堆肥初始期(A), 添加剂还没有对微生物产生显著影响, 3个处理丰度前15的属中有14种是相同的:乳杆菌属(Lactobacillus)、肠球菌属(Enterococcus)、维斯氏菌属(Weissella)、丹毒丝菌属(Erysipelothrix)、气球菌属(Aerococcus)、嗜冷杆菌属(Psychrobacter)、Jeotgalibaca、拟杆菌属(Bacteroides)、uncultured_bacterium_f_Aerococcaceae、赖氨酸芽胞杆菌属(Lysinibacillus)、棒状杆菌属(Corynebacterium_1)、不动杆菌(Acinetobacter)、鞘氨醇杆菌属(Sphingobacterium)和假单细胞菌(Pseudomonas), 变色杆菌(Paenochrobactrum)存在于CK和PG2处理中, 短状杆菌属(Brachybacterium)存在于BC2处理中.生物质炭改善了堆肥的孔隙及其通气状况, 创造了一个更有利于好氧短状杆菌(Brachybacterium)生长的环境(王海候等, 2020).
堆肥高温期(B), CK处理丰度前3的属为棒状杆菌属(Corynebacterium_1)(17.93%)、维斯氏菌属(15.89%)和鞘氨醇杆菌属(Sphingobacterium)(11.3%);BC2处理为棒状杆菌属(Corynebacterium_1)(20.58%)、维斯氏菌属(Weissella)(12.46%)和乳杆菌属(Lactobacillus)(11.36%);PG2处理为棒状杆菌属(Corynebacterium_1)(21.68%)、乳杆菌属(Lactobacillus)(18.6%)和维斯氏菌属(Weissella)(8.38%).生物质炭和凹凸棒的添加都增加了棒状杆菌属(Corynebacterium_1)、大洋芽胞杆菌属(Oceanobacillus)、嗜热菌属(Thermobifida)、Jeotgalicoccus的相对丰度, 降低了维斯氏菌属(Weissella)、不动杆菌属(Acinetobacter)和变色杆菌属(Paenochrobactrum)的相对丰度.生物质炭的添加更有利于Jeotgalicoccus的生长, 抑制不动杆菌属(Acinetobacter)的生长.凹凸棒的添加更有利于促进棒状杆菌属(Corynebacterium_1)、大洋芽胞杆菌属(Oceanobacillus)、嗜热菌属(Thermobifida)的生长, 抑制维斯氏菌属(Weissella)、变色杆菌属(Paenochrobactrum)的生长.棒状杆菌属(Corynebacterium_1)能够促进亚硝酸盐的产生, 对于烃类恶臭气体气体具有良好的降解作用(夏湘勤等, 2020).大洋芽胞杆菌属(Oceanobacillus)、嗜热菌属(Thermobifida)、全甘球菌(Jeotgalicoccus)都具有固氮能力(余震等, 2020;刘东明等, 2020;钱勋, 2016).维斯氏菌属(Weissella)和变色杆菌属(Paenochrobactrum)能促进反硝化作用(陆晓林等, 2019).不动杆菌(Acinetobacter)能够促进碳氮的代谢, 具有氮素转化的功能(李昌宁等, 2020).
堆肥降温期(C), CK处理丰度前3的菌属为芽孢杆菌属(Bacillus)(19.87%)、嗜热菌属(Thermobifida)(9.41%)和黄杆菌属(8.3%);BC2处理为芽孢杆菌属(Bacillus)(23.67%)、棒状杆菌属(Corynebacterium_1)(12.49%)和uncultured_bacterium_f_Sphingobacteriaceae(8.84%);PG2处理为芽孢杆菌属(Bacillus)(21.58%)、鞘氨醇杆菌属(Sphingobacterium)(13.19%)和uncultured_bacterium_f_Xanthomonadaceae(6.86%).芽孢杆菌属(Bacillus)成为3组处理的最优势菌属, 在CK处理中芽孢杆菌属(Bacillus)的相对丰度上升至19.87%, BC2和PG2处理分别上升至23.67%和21.58%, 生物质炭的添加更能够促进芽孢杆菌属(Bacillus)的生长.芽孢杆菌属(Bacillus)具有固氮能力(Xu et al., 2014), 对植物生长有促进作用(Park et al., 2019).生物质炭和凹凸棒的添加增加了纤细芽胞杆菌属(Gracilibacillus)和uncultured_bacterium_f_Xanthomonadaceae的相对丰度, 降低了乳杆菌属(Lactobacillus)和赖氨酸芽胞杆菌属(Lysinibacillus)的相对丰度.生物质炭的添加更好地抑制了乳杆菌属(Lactobacillus)的相对丰度, 凹凸棒的添加更有利于促进纤细芽胞杆菌属(Gracilibacillus)、uncultured_bacterium_f_Xanthomonadaceae的生长, 抑制赖氨酸芽胞杆菌属(Lysinibacillus)的生长.纤细芽胞杆菌属(Gracilibacillus)有固氮作用(陆晓林等, 2019).乳杆菌属(Lactobacillus)和赖氨酸芽胞杆菌(Lysinibacillus)是具有反硝化作用(王建才等, 2020).
堆肥腐熟期(D), CK处理丰度前3的菌属为鞘氨醇杆菌属(Sphingobacterium)(16.58%)、uncultured_bacterium_f_Sphingobacteriaceae(13.27%)和芽孢杆菌属(Bacillus)(12.87%);BC2处理为uncultured_bacterium_f_Sphingobacteriaceae(29.26%)、芽孢杆菌属(Bacillus)(15.7%)和鞘氨醇杆菌属(Sphingobacterium)(10.74%);PG2处理为鞘氨醇杆菌属(Sphingobacterium)(21.01%)、芽孢杆菌属(Bacillus)(14.46%)和黄杆菌属(Flavobacterium)(12.74%).生物质炭和凹凸棒的添加增加了uncultured_bacterium_f_Fodinicurvataceae和糖单孢菌属(Saccharomonospora)的相对丰度, 降低了肠球菌(Enterococcus)、气球菌属(Aerococcus)和极小单胞菌属(Pusillimonas)的相对丰度.生物质炭的添加更有利于uncultured_bacterium_f_Fodinicurvataceae和糖单孢菌属(Saccharomonospora)的生长, 而凹凸棒的添加更有利于抑制肠球菌(Enterococcus)、气球菌属(Aerococcus)和极小单胞菌属(Pusillimonas)的生长.糖单孢菌属(Saccharomonospora)具有降解木聚糖的能力, 有一定的固氮能力(鲍艳宇等, 2008).肠球菌属(Enterococcus)和极小单胞菌属(Pusillimonas)能够促进反硝化作用(邓雯文, 2019;Huang et al., 2019).气球菌属(Aerococcus)具有很强的耐盐能力并能够产酸, 同时能够降解有机物(唐伟欣等, 2020).
以上结果表明, 生物质炭的添加能够更好地促进Jeotgalicoccus、芽孢杆菌属(Bacillus)、uncultured_bacterium_f_Fodinicurvataceae和糖单孢菌属(Saccharomonospora)的生长, 抑制乳杆菌属(Lactobacillus)和不动杆菌属(Acinetobacter)的生长, 而添加凹凸棒更有利于促进棒状杆菌属(Corynebacterium_1)、大洋芽胞杆菌属(Oceanobacillus)、嗜热菌属(Thermobifida)、纤细芽胞杆菌属(Gracilibacillus)和uncultured_bacterium_f_Xanthomonadaceae的成长, 抑制维斯氏菌属(Weissella)、变色杆菌属(Paenochrobactrum)、赖氨酸芽胞杆菌属(Lysinibacillus)、肠球菌(Enterococcus)、气球菌属(Aerococcus)和极小单胞菌属(Pusillimonas)的生长.生物质炭的添加更能够促进相对丰度高于15%的优势固氮细菌芽孢杆菌属(Bacillus)的生长, 抑制相对丰度高于15%的优势反硝化细菌乳杆菌属(Lactobacillus)的生长.
3.6 优势菌群演替对碳氮损失的冗余分析冗余分析(RDA)是识别优势菌群和环境要素间相关性的重要手段(Li et al., 2017).图 6为堆肥过程中CK、BC2和PG2处理在不同阶段的碳氮损失与各时期丰度前15属之间的冗余分析(RDA)结果.结果表明, uncultured_bacterium_f_Fodinicurvataceae、极小单胞菌属(Pusillimonas)、芽孢杆菌属、Planifilum、Jeotgalicoccus、嗜热菌属(Thermobifida)、uncultured_bacterium_f_Xanthomonadaceae和糖单孢菌属(Saccharomonospora)与碳氮损失之间呈显著负相关, 表明这些微生物能减少堆肥过程中的碳氮损失.已有研究表明, 芽孢杆菌属(Bacillus)、嗜热菌属(Thermobifida)、糖单孢菌属(Saccharomonospora)具有固氮能力(Sangwan et al., 2004;黄翠, 2010;夏家帅, 2018).Planifilum能分泌木聚糖酶并具有固氮作用(Giri et al., 2007).极小单胞菌(Pusillimonas)具有反硝化功能(Huang et al., 2019).本研究结果表明, 堆肥添加剂可以显著增加以上固氮微生物的丰度, 降低反硝化菌极小单胞菌(Pusillimonas)的丰度, 从而增加堆肥中氮素的固定.生物质炭的添加能够更好地增加Jeotgalicoccus、芽孢杆菌属(Bacillus)、uncultured_bacterium_f_Fodinicurvataceae和糖单孢菌属(Saccharomonospora)的相对丰度, 添加凹凸棒可以更好地增加棒状杆菌属(Corynebacterium_1)、大洋芽胞杆菌属(Oceanobacillus)、嗜热菌属(Thermobifida)、纤细芽胞杆菌属(Gracilibacillus)和uncultured_bacterium_f_Xanthomonadaceae的相对丰度.堆肥过程中, 具有固氮功能细菌的增加和存在对堆肥质量的提高是有益的, 因此, 本结果可为堆肥中固氮微生物的筛选提供指导, 而筛选培养固氮微生物并应用到堆肥过程中, 对减少堆肥过程中养分的损失和提高堆肥质量具有重要意义.
 |
| 图 6 堆肥样品各处理碳氮损失及前15个属的冗余分析(RDA) Fig. 6 Redundancy analysis (RDA) of N-loss, C-loss and top 15 genera of each treatment of composting samples |
乳杆菌属(Lactobacillus)、肠球菌属(Enterococcus)、Paenochrobactrum、不动杆菌属(Acinetobacter)、气球菌属(Aerococcus)、维斯氏菌属(Weissella)、赖氨酸芽胞杆菌属赖氨酸芽胞杆菌(Lysinibacillus)、棒状杆菌属(Corynebacterium_1)与碳氮损失呈显著正相关, 表明这些细菌在堆肥过程的碳氮损失中起着重要作用.乳杆菌属(Lactobacillus)具有反硝化脱氮性能(Chèneby et al., 2000), 假单孢菌属(Pseudomonas)、肠球菌属(Enterococcus)和不动杆菌属(Acinetobacter)具有反硝化功能(Briones-Roblero et al., 2016;李一依等, 2020;王紫琪等, 2020), 魏斯氏菌属(Weissella)在发酵过程中产生一氧化氮, 是反硝化微生物(徐江兵等, 2017), 赖氨酸芽胞杆菌属(Lysinibacillus)对于强化反硝化过程具有潜在的积极作用(王紫琪等, 2020), 棒状杆菌属(Corynebacterium_1)具有反硝化作用(唐梦君, 2017).由此可见, 这些菌属都具有反硝化功能, 堆肥过程中应尽可能减少这些细菌的存在, 对减少堆肥过程中碳氮损失具有重要意义.本研究结果表明, 堆肥添加剂可以显著降低以上微生物的丰度, 从而减少堆肥中氮素的反硝化损失.而生物质炭的添加可以更好地降低乳杆菌属(Lactobacillus)和不动杆菌属(Acinetobacter)的相对丰度, 添加凹凸棒可以更好地降低维斯氏菌属(Weissella)、变色杆菌属(Paenochrobactrum)、赖氨酸芽胞杆菌属(Lysinibacillus)、肠球菌(Enterococcus)、气球菌属(Aerococcus)和极小单胞菌属(Pusillimonas)的相对丰度.
3.7 KEGG差异性分析KEGG数据库可以实现分析基因产物的功能及基因产物在细胞代谢中的作用(Minoru et al., 2007).图 7为堆肥过程中CK、BC2和PG2处理在第10 d的微生物群落功能的差异性分析(KEGG)结果.结果表明, 添加剂显著改变了优势微生物群落的功能基因.在众多功能基因中, 碳水化合物代谢(Carbohydrate metabolism)和氨基酸代谢(Amino acid metabolism)是微生物的重要生物代谢, 它们的代谢产物能够调控微生物群落结构和丰度变化(Guo et al., 2014), 并影响碳氮损失.CK、BC2和PG2处理的氨基酸代谢水平表现为CK>PG2>BC2(图 7a和7b), 碳水化合物代谢水平为BC2处理显著低于PG2和CK处理(图 7a和7c), PG2处理与CK处理之间碳水化合物代谢水平差异不显著.氨基酸作为细菌代谢的能量和碳源(López-González et al., 2015), 被认为是调节碳氮转化的主要影响因素(Lu et al., 2019), 而生物质炭可以改变碳水化合物中多糖的代谢过程(Soremi et al., 2018), 进而影响碳水化合物的代谢.本研究结果表明, 两种添加剂都显著抑制了堆肥中的氨基酸代谢, 且生物质炭的抑制作用大于凹凸棒.生物质炭的添加对碳水化合物代谢的抑制效果显著, 而凹凸棒对碳水化合物代谢的抑制效果不显著.由此可见, 生物质炭的添加能够更好地通过改变优势微生物群落及其功能基因而影响氨基酸和碳水化合物代谢, 从而减少堆肥过程中的碳氮损失.
 |
| 图 7 KEGG差异性分析 Fig. 7 KEGG difference analysis |
1) 不同添加量的生物质炭和凹凸棒都能够降低堆肥过程中的碳氮损失, 10%添加量的效果都优于5%添加量, 且生物质炭的效果优于凹凸棒.
2) 添加10%生物质炭和添加10%凹凸棒都能够显著促进堆肥中固氮类细菌的相对丰度, 而抑制反硝化细菌的相对丰度.生物质炭的添加更能够促进优势固氮微生物的成长, 抑制优势反硝化微生物的成长.
3) 添加10%生物质炭和添加10%凹凸棒都能够通过影响优势微生物群落及其氨基酸代谢和碳水化合物代谢功能基因抑制堆肥过程中的碳氮代谢, 从而减少碳氮损失, 且添加10%生物质炭对优势微生物群落及其氨基酸代谢和碳水化合物代谢功能基因影响更大.
Awasthi M K, Wang Q, Ren X, et al. 2016. Role of biochar amendment in mitigation of nitrogen loss and greenhouse gas emission during sewage sludge composting[J]. Bioresource Technology, 219: 270-280. DOI:10.1016/j.biortech.2016.07.128 |
Bernal M P, Alburquerque J A, Moral R. 2009. Composting of animal manures and chemical criteria for compost maturity assessment[J]. Bioresource technology, 100: 5443-5444. |
Briones-Roblero C I, Rodriguez-Diaz R, Santiago-Cruz J A, et al. 2016. .Deradation capacities of bacteria and yeasts isolated from the gut of Dendroctonus rhizophagus[J]. Folia Microbiplogica, 62(1): 1-9. |
鲍艳宇, 周启星, 颜丽, 等. 2008. 不同畜禽粪便堆肥过程中有机氮形态的动态变化[J]. 环境科学学报, 28(5): 930-936. DOI:10.3321/j.issn:0253-2468.2008.05.019 |
Chèneby D, Philippot L. 2000. 16S rDNA analysis for characterization of denitrifying bacteria isolated fromthree agricultural soils[J]. FEMS Microbiology Ecology, 34: 121-128. DOI:10.1111/j.1574-6941.2000.tb00761.x |
邓雯文, 陈姝娟, 何雪萍, 等. 2019. 鸡粪-堆肥中重金属残留、抗生素耐药基因及细菌群落变化研究[J]. 农业环境科学学报, 38(2): 439-450. |
Giri S, Pati B R. 2004. A comparative study on phyllosphere nitrogen fixation by newly isolated Corynebacterium sp.& Flavobacterium sp. and their potentialities as biofertilizer[J]. Acta Microbiology Immunol Hung, 51(1/2): 47-56. |
Gul S, Whalen J, Kthomas B W, et al. 2015. Physico-chemical properties and microbial responses in biochar-amended soils: mechanisms and future directions[J]. Agric Ecosyst Environment, 206: 46-59
|
Gao M, Liang F, An Y, et al. 2015. Evaluation of stability and maturity during forced-aeration composting of chicken manure and sawdust at different C/N ratios[J]. Chemosphere, 78(5): 614-619. |
Guo X, Liu S, Wang Z, et al. 2014. Metagenomic profiles and antibiotic resistance genes in gut microbiota of mice exposed to arsenic and ron[J]. Chemosphere, 112: 1-8. DOI:10.1016/j.chemosphere.2014.03.068 |
郭梅莹. 2015. 凹凸棒添加堆肥过程中的酶活性研究[D]. 兰州: 兰州交通大学
|
黄华, 黄懿梅, 刘学玲, 等. 2013. 覆盖处理对猪粪秸秆堆肥中氮素转化和堆肥质量的影响[J]. 环境科学学报, 33(3): 780-786. |
贺琪, 李国学, 张亚宁, 等. 2005. 高温堆肥过程中的氮素损失及其变化规律[J]. 农业环境科学学报, (1): 169-173. DOI:10.3321/j.issn:1672-2043.2005.01.037 |
Hong N, Chen J, Gao D, et al. 2014. Enhanced water reduction by turning during sewage sludge composting[J]. Journal of Chemical Technology&Biotechnology, 89(5): 9-13. |
黄翠, 杨朝晖, 肖勇, 等. 2010. 堆肥嗜热纤维素分解菌的筛选鉴定及其强化堆肥研究[J]. 环境科学学报, 30(12): 2457-2463. |
Huang G F, Wong J. 2004. Effect of C/N on composting of pig manure with sawdust[J]. Waste Manage, 24(8): 805-813. DOI:10.1016/j.wasman.2004.03.011 |
IBI(Internationa Biochar Initiative). 2012. Standardized product definition and product testing guidelines for biochar that is used in soil[Z]. IBI-STD-01
|
李波, 叶菁, 刘岑薇, 等. 2017. 生物炭添加对猪粪堆肥过程碳素转化与损失的影响[J]. 环境科学学报, 37(9): 3511-3518. |
李昌宁, 苏明, 姚拓, 等. 2020. 微生物菌剂对猪粪堆肥过程中堆肥理化性质和优势细菌群落的影响[J]. 植物营养与肥料学报, 26(9): 1600-1611. |
刘东明, 孟繁华, 郝艳, 等. 2020. 微生物菌剂对堆肥功能微生物重构的影响[J]. 环境科学研究, 33(9): 2011-2019. |
李丹阳, 马若男, 亓传仁, 等. 2020. 含水率对羊粪堆肥腐熟度及污染气体排放的影响[J]. 农业工程学报, 36(20): 254-262. DOI:10.11975/j.issn.1002-6819.2020.20.030 |
Lei F, Vandergheynst J S. 2000. The effect of microbial inoculation and pH on microbial community structure changes during composting[J]. Process Biochemistry, 35(9): 923-929. DOI:10.1016/S0032-9592(99)00155-7 |
李国学, 李玉春, 李彦富. 2003. 固体废物堆肥化及堆肥添加剂研究进展[J]. 农业环境科学学报, (2): 252-256. DOI:10.3321/j.issn:1672-2043.2003.02.034 |
Li Q, Guo X, Lu Y, et al. 2016. Impacts of adding FGDG on the abundance of nitrification and denitrification functional genes during dairy manure and sugarcane pressmudco-composting[J]. Waste Management, 56: 63-70. DOI:10.1016/j.wasman.2016.07.007 |
Li H, Duan M, Gu J, et al. 2017. Effects of bamboo charcoal on antibiotic resistance genes during chicken manure composting[J]. Ecotoxicol Environment Safety, 140: 1-6. DOI:10.1016/j.ecoenv.2017.01.007 |
李荣华, 涂志能, Ali Amjad, 等. 2020. 生物炭复合菌剂促进堆肥腐熟及氮磷保留[J]. 中国环境科学, 40(8): 3449-3457. DOI:10.3969/j.issn.1000-6923.2020.08.024 |
Luo Y, Yu Z Y, Zhang K L, et al. 2016. The properties and functions of biochars in forest ecosystems[J]. Soils Sediments, 16: 2005-2020. DOI:10.1007/s11368-016-1483-5 |
López-González J, Suárez-Estrella F, Vargas-García M, et al. 2015. Dynamics of bacterial microbiota during lignocellulosic waste composting: Studies upon its structure, functionality and biodiversity[J]. Bioresource Technology, 175: 406-146. DOI:10.1016/j.biortech.2014.10.123 |
Lu Y, Zheng G, Zhou W, et al. 2015. Bioleaching conditioning increased the bioavailability of polycyclic aromatic hydrocarbons to promote their removal during co-composting of industrial and municipal sewage sludges[J]. Science of the Total Environmental, 665: 1073-1082. |
陆晓林, 戴传超, 贾永, 等. 2019. 堆肥过程中优势功能微生物菌群多样性及其影响因素[J]. 生态学杂志, 38(9): 2866-2874. |
林小凤, 李国学, 任丽梅, 等. 2008. 氯化铁和过磷酸钙控制堆肥氮素损失的效果研究[J]. 农业环境科学学报, (4): 1662-1666. DOI:10.3321/j.issn:1672-2043.2008.04.067 |
李一依, 赵勇娇, 刘芳, 等. 2020. 马赛菌属Massilia neuiana异养硝化-好氧反硝化脱氮特征实验[J]. 环境工程, 38(10): 103-107. |
Minoru K, Michihiro A, Susumu G, et al. 2008. KEGG for linking genomes to life and the environment[J]. Nucleic Acids Research, 36(1): 480-484. |
Nelissen V, Rutting T, Huygens D, et al. 2012. Maize biochars accelerate short-term soil nitrogen dynamics in a loamy sand soil[J]. Soil Biology and Biochemistry, 55: 20-27. DOI:10.1016/j.soilbio.2012.05.019 |
Pan J, Li R, Zhai L, et al. 2019. Influence of palygorskite addition on biosolids composting process enhancement[J]. Journal of Cleaner Production, 217: 371-379. DOI:10.1016/j.jclepro.2019.01.227 |
Park G, Nam J, Kim J, et al. 2019. Structure and mechanism of surfactin peptide from Bacillus velezensis antagonistic to fungi plant pathogens[J]. Bulletin of the Korean Chemical Society, 40(7): 704-709. DOI:10.1002/bkcs.11757 |
钱勋. 2016. 好氧堆肥对畜禽粪便中抗生素抗性基因的削减条件探索及影响机理研究[D]. 咸阳: 西北农林科技大学
|
Sangwan P, Chen X, Hugenholtz P, et al. 2004. Janssen pH.Chthoniobacter flavus gen.nov, sp.nov., the First pure-culture representative of subdivision two, Spartobacteria classis nov., of the Phylum Verrucomicrobia[J]. Applied & Environmental Microbiology, 70(10): 5875-5881. |
Soremi A O, Adetunji M T, Azeez J O, et al. 2018. Effects of soil amendment on potassium fractions of southwestern nigerian soils[J]. Communications in Soil Science and Plant Analysis, 49(10): 1186-1198. DOI:10.1080/00103624.2018.1455847 |
唐梦君. 2017. 一株嗜麦芽寡养单胞菌的筛选鉴定及其聚磷和降解有机磷的研究[D]. 武汉: 湖北大学
|
唐伟欣, 孙兴滨, 高浩泽, 等. 2020. 规模化畜禽养殖场粪便中多重耐药菌分离鉴定及其耐药特征[J]. 农业环境科学学报, 39(1): 207-216. |
吴梦婷, 梅娟, 苏良湖, 等. 2020. 硫酸亚铁和过磷酸钙对牛粪秸秆混合堆肥氮损失和腐殖化的影响[J]. 生态与农村环境学报, 36(10): 1353-1361. |
Wang P, Chang C M, Watson M E, et al. 2004. Maturity indices for composted dairy and pig manures[J]. Soil biology and biochemistry, 36(5): 767-776. DOI:10.1016/j.soilbio.2003.12.012 |
王紫琪, 杨基先, 何伟华, 等. 2020. 一株异养硝化-好氧反硝化菌Bacillus flexus X1-L的分离鉴定及效能研究[J]. 环境科学学报, 40(10): 3742-3748. |
王海候, 韩奇隆, 刘泽凯, 等. 2020. 不同热解温度生物质炭对羊粪堆肥过程氮素损失的减控效果[J]. 农业工程学报, 36(19): 184-191. DOI:10.11975/j.issn.1002-6819.2020.19.021 |
王建才, 朱荣生, 刘兴华, 等. 2020. 微生物菌剂对猪粪堆肥中细菌群落结构的影响[J]. 应用生态学报, 31(7): 2449-2456. |
夏家帅. 2018. 柑橘皮渣堆肥工艺优化及其微生物群落研究[D]. 重庆: 重庆大学
|
夏湘勤, 唐朱睿, 黄彩红, 等. 2020. 环丙沙星胁迫猪粪生物转化中抗性基因微生物响应分析[J]. 环境科学研究, 33(7): 1711-1720. |
Xu L H, Zeng X C, Nie Y, et al. 2014. Pontibacter diazotrophicus sp.nov., anovel nitrogen fixing bacterium of the family Cytophagaceae[J]. PLoS One, 9(3): 1-7. |
徐江兵, 王艳玲, 罗小三, 等. 2017. 纳米氧化锌对堆肥过程中细菌群落演替的影响[J]. 应用与环境生物学报, 23(6): 1166-1171. |
殷亚楠. 2018. 铜对畜禽粪便堆肥过程中微生物群落及抗性基因影响机理研究[D]. 咸阳: 西北农林科技大学
|
余震, 周顺桂. 2020. 超高温好氧发酵技术: 堆肥快速腐熟与污染控制机制[J]. 南京农业大学学报, 43(5): 781-789. |
张嘉超. 2009. 农业废物好氧堆肥过程因子对细菌群落结构的影响[D]. 长沙: 湖南大学
|