 2021, Vol. 41
2021, Vol. 41



2. 杭州环境集团有限公司, 杭州 310022
2. Hangzhou Environmental Group Company Limited, Hangzhou 310022
全球每年接近13~16亿t的食物, 如新鲜蔬菜、水果、面包、奶制品等在食物供应链中被浪费.餐厨垃圾占据了每年食物总产量的1/3, 且此比例每年仍在上升, 造成了严重的资源浪费和环境污染(Braguglia et al., 2018).在中国, 餐厨垃圾含水率高达60%~90%, 可降解的部分比例约为41%~62%碳水化合物, 15%~25%蛋白质, 和13%~30%油脂(Fisgativa et al., 2016).为了降低餐厨垃圾的环境危害和能源浪费, 基于餐厨垃圾高水分、低热值、不稳定的特性, 传统的餐厨垃圾处理方式包括焚烧、填埋、堆肥、热处理以及用作动物饲料(Li et al., 2018a).现阶段, 餐厨垃圾发酵处理已经被证明是一种可以同时降低餐厨垃圾环境危害并可回收能量的有效的处理方式(Yun et al., 2018).关于餐厨垃圾发酵产氢产甲烷已经有很多研究(Cheng et al., 2016a; De Gioannis et al., 2017; Xiao et al., 2018).这些发酵方式一定程度上增加了甲烷的产率, 提升了整体的能量转化效率.然而餐厨垃圾中油脂含量高, 油脂降解慢且容易包裹微生物等问题十分严重.尤其中国饮食习惯造成的餐厨垃圾中油脂成分含量很高, 经过机械法除油后仍然有大量油脂进入发酵体系, 容易造成发酵体系崩溃.预处理是一种降低油脂发酵抑制和提升发酵性能的一种有效的方式.
餐厨垃圾的预处理方式可以分为加热处理、机械处理(微波、超声波、高压脉冲放电等)、生物处理(酶水解和好氧堆肥等)、化学处理(加酸、碱和氧化物)4大类(Bong et al., 2018).关于餐厨垃圾生物质预处理方式对于发酵氢气和甲烷产率的提高主要体现于增加了溶解性还原糖的含量(Jarunglumlert et al., 2018).然而对于难于降解的餐厨垃圾中的蛋白质和油脂成分发酵产氢和产甲烷却几乎没有涉及, 尤其是油脂成分的发酵更是一种难以处理的问题.而目前关于油脂预处理用于厌氧发酵的研究较少.Zhang等研究了餐厨垃圾和粗油不同比例混合厌氧发酵产甲烷的效果(Zhang et al., 2017b).当8~10 g·L-1的餐厨添加30%的浮油时, 甲烷产率最高, 达到573.0 mL·g-1.浮油超过35%时, 由于pH的降低和大量脂肪酸的积累, 甲烷产率大幅降低.Liu等提取了一株Klebsiella pneumoniae sp. LZU10菌群, 用于预处理降解油脂.油脂降解率可以达到39.5%~59.6%, 甲烷产率提高了41% (Liu et al., 2018).然而生物预处理步骤较为繁琐, 而处理时间较长.Meng的研究发现, Lipase系列3种酶用于处理动物油脂和植物油脂, 在条件为24 h, 1000~1500 μL和40~50 ℃时, 处理效果最好, 整体甲烷产率分别提升了80.8%~157.7%、26.9%~53.8%和37.0%~40.7% (Meng et al., 2015).生物酶处理是一种可行的油脂预处理方式, 然而酶处理的成本高, 工业化程度较低, 一个可行的例子是脂肪酶可用于废纸造纸的油脂去除, 然而在废弃物厌氧发酵领域, 酶的应用程度不高.
而本文涉及的预处理方式为水热和臭氧的预处理.水热预处理改变了底物的物理化学特征, 而较高温度和较长时间的预处理对于发酵的影响主要体现在溶解COD及高温高压产生的高能基团.Ding发现, 140 ℃和20 min的餐厨垃圾经过水热预处理后, 两阶段氢气甲烷产率分别达到峰值, 为43.0和511.6 mL·g-1 (Ding et al., 2017b).臭氧由于其具有活性的高能氧原子, 常用于处理化学需氧量(COD)含量高的废水和固废等废弃物处理.在已有的研究中, 水热(Cheng et al., 2020)预处理以及臭氧(Yue et al., 2020)预处理已经被验证可以促进油脂中常见组分甘油三油酸酯的水解和降解, 并且可以促进厌氧发酵产甲烷或者与餐厨垃圾混合厌氧发酵产甲烷, 缓解油脂包裹微生物的问题, 然而尚且缺少不同组分实际油脂的预处理及厌氧发酵产甲烷效果验证, 也尚无水热以及臭氧这两种可以产生高能自由基的预处理方式的效果比较.
在这项研究中, 预处理前后的废弃油脂微观结构及官能团变化可以通过扫描电子显微镜(SEM)、以及傅里叶转换红外线光谱分析仪(FTIR)进行测定.本研究的目标为:①研究水热及臭氧预处理对于油脂降解的效果和底物特性的影响;②研究水热及臭氧预处理对于废弃油脂厌氧发酵产甲烷效果的影响;③通过预处理后暗发酵产甲烷的方式, 提升甲烷产率以及整体能量转化效率.
2 实验材料与方法(Materials and methods) 2.1 发酵原料和特性厌氧发酵使用的油脂有3种, 分别为中国餐厨垃圾中的典型油脂:火锅油、烧烤油和川菜油.实验所用的油脂分别来源于浙江省杭州市的某火锅店、烧烤店以及川菜馆.油脂经过过滤、除杂后存储于4 ℃条件下.油脂的成分如表 1所示.3种油脂的成分各不相同, 用于研究预处理方式对于不同成分油脂降解以及厌氧发酵的影响.水分含量通过在105 ℃烘箱内烘烤至质量不变来测定, 有机质及灰分含量通过在450 ℃马弗炉内灼烧2 h来测定(Lin et al., 2017).餐厨垃圾中的还原糖含量通过DNS法进行测试, 蛋白质和油脂成分分别采用Lowry以及萃取方法(Cheng et al., 2019b)进行测试.工业分析, 元素分析以及热值通过之前的测试方法进行测试(Cheng et al., 2019a).预处理后油脂固体底物的微观结构的测试方法包括SEM(台式显微镜TM-1000, HITACHI)及FTIR (气相色谱仪——傅里叶红外光谱仪SGE/Agilent 6890/Nicolet 5700).
| 表 1 油脂底物的特性 Table 1 Characterization of lipid feedstocks |
3种废弃油脂生物质在发酵前, 采用的是水热或臭氧预处理方式.5 g底物油脂和100 mL的稀氢氧化钠溶液(2%V/V)加入水热反应釜(Parr 4500, 美国)中, 密闭容器加热至220 ℃恒温并保持30 min.在臭氧预处理过程中, 使用的是纯氧源臭氧发生器(ECON, 中国).5 g的底物油脂加入发酵瓶中, 加入去离子水使总体积为100 mL.使用1 mol·L-1的NaOH溶液调节pH为8.0.产生的臭氧通过长钢管道(180 mm × Φ 3 mm)导入417 mL的发酵瓶, 恒定流速为1 L·min-1.该实验的臭氧浓度设置为0.2 g-O3/g-TVS.此臭氧投加量源于已有研究中甘油三油酸酯臭氧投加量的最佳量(Yue et al., 2020), 并且预实验也体现了相类似的规律.
废弃火锅油脂、烧烤油脂和川菜油脂的主要油脂成分为甘油三酯, 包括甘油三油酸酯、甘油三硬脂酸酯、甘油三十六酸酯等.理论上1 mol甘油三酯大分子可以水解为1 mol甘油和3 mol大分子羧酸, 而水解出的甘油和羧酸部分可以用于发酵产生甲烷(Xia et al., 2014).以甘油三油酸酯为例, 相关的水解和反应的原理式如式(1)所示(Dong et al., 2009).
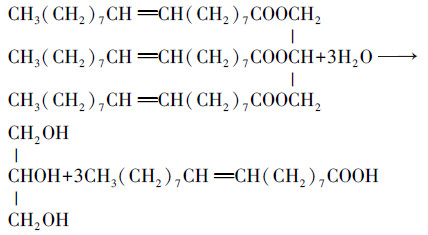
|
(1) |
长链脂肪酸(如油酸、硬脂酸等)通过β-氧化途径被逐步降解利用(Adeva-Andany et al., 2019), 利用的方式见式(2).
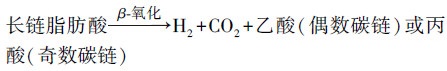
|
(2) |
通过β-氧化的过程, 长链脂肪酸每次脱下2个碳原子, 参与到产甲烷古菌的代谢过程中, 形成各种代谢产物.具有偶数碳链的长链脂肪酸可以完全降解, 而具有奇数碳链的长链脂肪酸最后剩余3个碳原子形成丙酸.在产甲烷过程乙酸和丁酸均可以作为底物被产甲烷菌利用产甲烷.
2.3 产甲烷接种物发酵实验所用的产甲烷菌分离自中国浙江省杭州市杭州环境集团甲烷发酵罐, 取分离后的500 mL活性污泥于1 L锥形瓶中, 在厌氧工作站(Whitley DG250, 美国)中放置30 d消耗完原有有机质.之后将pH调整为8.0±0.1以抑制产氢细菌等杂菌.瓶中加入1.25 g纤维素, 置于厌氧工作站中富集培养.14 d后即得到富集完成的产甲烷菌.经过16S rRNA测定, 产甲烷菌的主要菌种为Methanosarcina和Methanothrix (Cheng et al., 2011).
2.4 发酵产甲烷方法批次发酵产甲烷的实验在417 mL规格的发酵瓶中进行.3组实验分别采用三重样.取100 mL预处理之后的浊液(对应5 g原始底物), 利用6 mol·L-1的盐酸溶液和6 mol·L-1的氢氧化钠溶液将发酵系统的pH调节为8.0±0.1, 之后按照底物TVS:产甲烷菌活性污泥TVS=1∶2(体积比)接种产甲烷菌污泥, 加入灭菌后的去离子水混合使总体积为300 mL, 随后重新调节pH值为8.0±0.1.发酵瓶使用橡胶塞密封并以40 mL·min-1的流量通氮气10 min, 随后置于恒温水浴锅中进行厌氧发酵, 与1000 mL有机玻璃集气槽连接用于收集产生的气体.恒温水浴温度保持为(37.0±1.0) ℃.
2.5 分析测试方法厌氧发酵产甲烷过程产生的气体主要为CO2和CH4两种, 还混杂了N2、O2以及水蒸气、H2S等杂质气体.生物气成分利用带有热导检测器(TCD)的Agilent 7820A气相色谱仪进行测定.产气的总体积通过集气槽读数与发酵瓶顶部空间气体相加得到.甲烷发酵液中所含有的液相代谢产物(SMPs)的测定同样采用气相色谱法, 实验室所用的是同样是带有氢离子火焰检测器(FID)的气相色谱仪(型号:Agilent 7820A).
实验过程中得到的甲烷和二氧化碳的产量是根据实际发酵生物气的产量以及甲烷和二氧化碳的浓度分别计算, 并且根据标准温度和压力校正得到.甲烷产率是甲烷产量(mL)和底物的有机质干重(g)的比值(Ding et al., 2017a).能量转化效率定义为产生甲烷的能量与底物的能量之比(Ding et al., 2017a).
3 结果与讨论(Results and discussion) 3.1 废弃油脂的预处理降解水热预处理和臭氧预处理的高能自由基可以冲击大分子羧酸的化学键, 使大分子羧酸中的大分子C原子数降低.低C原子数的羧酸则更容易被产甲烷菌利用(Kallscheuer et al., 2017).有研究发现, 经过臭氧预处理之后, 甘油三油酸酯的主要羧酸成分为没有分解的油酸, 脱掉两个C原子的十六酸以及脱掉4个C原子的十四酸.在臭氧和碱(pH约为8.0)的作用下, 高能自由基诱导C18通过β氧化作用逐步降解为C16和C14.而臭氧浓度对于臭氧碱预处理的效果有明显的影响.臭氧浓度越高, 未降解的油酸的比例越低, 降解得到的C16和C14的比例越高.臭氧浓度越高, 高能自由基诱导C18通过β氧化作用反应速率越快, 反应越彻底(Yue et al., 2020).而油脂水热预处理的机制可以通过以下方面来说明.经过水热预处理之后, 甘油三油酸酯的主要成分为没有分解的甘油三油酸酯, 脱掉一个油酸的甘油二油酸酯, 脱掉两个油酸的甘油单油酸酯, 油酸以及十六酸.而水相中的主要成分为脂肪酸钠和甘油.水热加碱预处理的作用主要包括两个.①水解.在碱的作用下, 甘油三油酸酯逐渐水解为甘油和脂肪酸.②降解.在水热和碱的作用下, 高能自由基以及高压的作用下通过β氧化作用C18降解为C16和CO2 (Cheng et al., 2020).
经过水热和臭氧预处理后废弃火锅油脂混合物的FTIR谱图如图 1a所示, FTIR谱图可以显示两种预处理对于底物的组分与化学结构的影响.从图中进行分析, 预处理前后含有的化学官能团基本相同.3种底物在波长为2930、2850、1755、1550、1430以及1138 cm-1的峰可以识别.3种样品在1430 cm-1以及1138 cm-1出现了特征吸收峰, 这两个峰分别是—CH2的剪切振动峰和O—H伸缩振动峰(Francioso et al., 2010).两种样品在2930 cm-1处的特征吸收峰是—CH2的伸缩振动峰, 样品在2850 cm-1处的特征吸收峰是—CH的伸缩振动峰, 这两个峰是油脂或脂肪酸的碳链的特征峰(Francioso et al., 2010), 样品在1550 cm-1处的特征吸收峰是羧酸CO官能团的伸缩振动峰, 这个峰是有机酸类化合物的特征峰, 样品在1755 cm-1处的特征吸收峰是酯类CO官能团的伸缩振动峰, 这个峰是酯类化合物的特征峰(Francioso et al., 2010).与没有经过预处理的样品对比, 经过水热或臭氧预处理实验组的样品在1755 cm-1处吸收峰强度明显减弱, 而在1550 cm-1处吸收峰强度明显增强.从谱图中可以看出, 预处理实验组的实验样品和没有经过预处理实验组的实验样品相比, 酯类含量降低, 羧酸类含量上升.没有经过预处理的废弃油脂由于分子量很大, 难以直接被产甲烷菌利用, 在甲烷菌的量相同的情况下厌氧发酵处理能力会降低, 而水热和臭氧预处理将大分子油脂降解为相对小分子的羧酸, 降低了菌群对底物的利用难度.预处理效果可以用实际产甲烷效果进行验证.
 |
| 图 1 火锅油脂臭氧/水热预处理后FTIR谱图和SEM图像 (a.FTIR谱图, b.未预处理油脂SEM图像, c.臭氧预处理油脂SEM图像, d. 水热预处理油脂SEM图像) Fig. 1 FTIR spectra and SEM images of hotpot lipids after pretreatment. (a. FTIR spectra, b. SEM image without pretreatment, c. SEM image with ozonation pretreatment, d. SEM image with hydrothermal pretreatment) |
通过SEM分析油脂生物质的表面形态如图 1b~1d所示.预处理前, 油脂生物质呈不规则块状(10~50 μm), 表面光滑且存在褶皱.块状颗粒体积较大, 且内部致密.这是由于没有经过预处理的火锅油脂生物质在常见发酵温度(37 ~55 ℃)呈现为固体且积聚, 因而表现为大块固体形状.进过水热和臭氧预处理之后, 形成了大量1~5 μm左右的碎片, 而团块表面则存在大量孔隙, 孔隙直径为0.2~1 μm左右.这是由于经过臭氧或水热预处理之后, 油脂生物质成为蓬松结构, 大分子甘油三酯脱出甘油, 降解出大量的脂肪酸.团块之间的孔隙也会变多而且变小.然而水热的预处理效果不如臭氧预处理效果.这可能是因为臭氧预处理的过程中会产生大量的高能氧原子, 较之水热预处理的反应过程更为剧烈.因而在两种预处理条件下, 臭氧预处理可以导致更多的生物质碎片和更多孔隙的形成.
3.2 预处理后3种废弃油脂厌氧发酵产甲烷3种废弃油脂厌氧发酵甲烷的产率以及产甲烷速率如图 2所示.在经过48 d左右, 发酵产甲烷量不再发生明显变化, 可以认为产甲烷阶段结束.没有经过预处理的火锅油脂甲烷产率仅为(727.51±23.69) mL·g-1, 在第24 d达到产甲烷速率峰值(31.13±2.18) mL·g-1·d-1.经过水热预处理的火锅油脂甲烷产率提升至(779.72±22.12) mL·g-1, 在第20 d达到产甲烷速率峰值(93.73±2.50) mL·g-1·d-1.甲烷产率较之未处理组提升约7.2%, 而达到峰值速率的时间缩短了4 d.经过臭氧预处理的火锅油脂甲烷产率提升至(854.20±10.28) mL·g-1, 在第20 d达到产甲烷速率峰值(122.06±3.46) mL·g-1·d-1.甲烷产率较之未处理组提升约17.4%, 而达到峰值速率的时间缩短了4 d.可以发现在速率曲线中出现了一个峰, 这个峰是长链脂肪酸通过β-氧化过程逐渐降解为短链脂肪酸并且被产甲烷菌利用产甲烷的峰.没有经过臭氧或水热预处理的实验组的β-氧化过程缓慢.甲烷产率和产甲烷峰值速率也较低.这和SEM的测试结果是相符的.经过臭氧或水热预处理之后, 油脂生物质成为蓬松结构, 大分子甘油三酯脱出甘油, 降解出大量的脂肪酸, 团块之间的孔隙也会变多而且变小, 底物的利用增强.大分子酸的浓度过高会对甲烷菌的代谢过程造成阻碍, 甚至会抑制甲烷菌的活性.经过预处理后, 大分子甘油三酸酯被降解, 降低了甲烷菌对于底物的利用难度.同样, 这和FTIR测试结果也是相符的.没有经过预处理的废弃油脂由于分子量很大, 难以直接被产甲烷菌利用, 在甲烷菌的量相同的情况下厌氧发酵处理能力会降低, 而水热和臭氧预处理将大分子油脂降解为相对小分子的羧酸, 降低了菌群对底物的利用难度.因而产甲烷峰值速率提高, 甲烷产率提升, 达到甲烷峰值速率的时间也逐渐缩短.烧烤油和川菜油的厌氧发酵产甲烷结果也呈现了相同的规律.臭氧预处理后烧烤油的甲烷产率((771.85±5.18) mL·g-1)远高于没有经过预处理的烧烤油甲烷产率((693.69±9.88) mL·g-1), 而达到峰值速率的时间也由24 d缩短至20 d.臭氧预处理后川菜油的甲烷产率(801.69±9.58 mL·g-1)远高于没有经过预处理的烧烤油甲烷产率((734.04±11.22) mL·g-1), 而达到峰值速率的时间也由20 d缩短至16 d.然而3种废弃油脂甲烷产率、产甲烷速率以及达到峰值速率的时间也略有不同.这是3种废弃油脂成分不同导致的.废弃火锅油脂中油脂的含量最高, 而较之于相同质量的蛋白质和糖类, 发酵产甲烷的潜力最强.然而油脂的降解则较慢, 因而达到峰值速率的时间也有所延长.因而以火锅油脂为例进行下列的分析.
 |
| 图 2 3种废弃油脂臭氧/水热预处理后产甲烷效果对比 (a.火锅油脂产甲烷速率, b.火锅油脂甲烷产率, c.烧烤油脂产甲烷速率, d.烧烤油脂甲烷产率, e.川菜油脂产甲烷速率, f.川菜油脂甲烷产率) Fig. 2 Methane production from pretreated waste lipids during anaerobic digestion. (a. methane production rate from hotpot lipids, b. methane yield from hotpot lipids, c. methane production rate from barbecue lipids, d. methane yield from barbecue lipids, e. methane production rate from Sichuan cuisine lipids, f. methane yield from Sichuan cuisine lipids) |
臭氧和水热预处理废弃火锅油脂发酵产甲烷过程的发酵液的sCOD的变化测试结果如图 3a所示.未经预处理的油脂发酵液sCOD开始为(740±80) mg·L-1.随着油脂被逐渐降解为小分子酸, sCOD浓度逐渐升高至(4040±140) mg·L-1并缓慢下降, 48 d时趋近于平缓, 厌氧发酵结束时sCOD较高, 约为2500 mg·L-1.这可能是由于没有经过预处理的油脂降解过程缓慢, 发酵过程中大量产甲烷菌群被油脂包裹.液相及油相组分均难以被甲烷菌利用, 使sCOD逐渐升高, 从而导致了甲烷产率和甲烷峰值速率的降低.而臭氧及水热组的发酵液初始sCOD略微高于单产组的sCOD.这可能是因为预处理过程产生的小分子甘油和脂肪酸等产物提高了sCOD.对于臭氧预处理组, 发酵液sCOD起始为(1620±210) mg·L-1.对于水热联产组, 发酵液sCOD起始为(1300±90) mg·L-1.随着发酵产甲烷过程的进行, 预处理组发酵液的sCOD均逐渐上升至大约5000 mg·L-1.发酵产甲烷过程结束后sCOD降低至低于1100 mg·L-1.这是因为在油脂被甲烷菌利用的过程中, 底物逐渐降解得到的小分子被产甲烷菌有效利用, 几乎不会积累, 因而sCOD逐渐降低.这和发酵产甲烷的结果是相符合的.
 |
| 图 3 火锅油脂臭氧/水热预处理后产甲烷过程sCOD和总SMPs变化 (a. sCOD浓度变化, b.总SMPs浓度变化, c.乙酸浓度变化, d.丁酸浓度变化) Fig. 3 sCOD concentration and total SMPs concentration during anaerobic digestion. (a. sCOD concentration, b. total SMPs concentration, c. acetate concentration, d. butyrate concentration) |
臭氧和水热废弃火锅油脂发酵产甲烷过程的发酵液的SMPs的变化测试结果如图 3b所示.发酵过程测试所得SMPs的主要成分为乙酸和丁酸, 此外还有少量的丙酸、异丁酸、异戊酸、戊酸和己酸, 这和甲烷古菌的发酵代谢途径和对于小分子酸的利用途径是相符的(Xia et al., 2015).油脂没有经过预处理厌氧发酵产甲烷过程中, 总SMPs由最开始的(5.39±1.09) mmol·L-1逐渐上升, 在30 d达到峰值(203.33±22.16) mmol·L-1, 之后逐渐下降.发酵结束48 d时SMPs浓度为(46.05±8.06) mmol·L-1.而经过臭氧或水热预处理后, SMPS峰值则分别上升至(240.31±8.97) mmol·L-1以及(220.40±20.26) mmol·L-1, 而达到峰值的时间分别缩短至24 d.48 d发酵结束时SMPs浓度低于25 mmol·L-1.这是由于没有经过水解和降解的油脂的主要成分甘油三酸酯分子量很大, 大分子的油脂难以被产甲烷菌直接利用而β-氧化过程缓慢.因而经过预处理之后, 水解和降解效果增强, 甘油部分被利用程度增强, SMPs浓度越高, 油脂产甲烷过程的SMPs的浓度有所提升, 达到峰值的时间越短.同样, 乙酸和丁酸的变化过程图 3c以及图 3d呈现了相似的规律.值得注意的是, 发酵结束后未处理的油脂组仍然残留大量的SMPs, 这也说明臭氧或水热预处理可以在一定程度上促进油酸的降解, 并且提高产甲烷菌对底物的利用从而提高发酵产甲烷的效率, 这与发酵产甲烷的结果是一致的.
3.3 3种废弃油脂厌氧发酵产甲烷能量转化效率3种油脂经过不同处理方式后用于厌氧发酵产甲烷的能量转化效率如图 4所示.能量转化效率按照甲烷热值801 kJ·mol-1, 以及如表 1所示的油脂热值进行计算.3种油脂能量转化效率呈现相似的规律, 即经过臭氧预处理油脂的厌氧发酵产甲烷能量转化效率最高, 没有经过预处理油脂的产甲烷效率最低.预处理对于油脂能量转化效率提升最高的为火锅油脂, 由没有经过预处理的64.88%提升到臭氧预处理后的76.18%, 提升比例约为17.42%.臭氧和水热预处理可以在一定程度上促进油脂的降解, 并且提高产甲烷菌对油脂底物的利用, 从而提高厌氧发酵产甲烷的效率.以火锅油脂的预处理厌氧发酵为例讨论完能量的输入和产出平衡问题.经过计算, 水热预处理的能量消耗为9.15 kJ·g-1, 而臭氧预处理的能量消耗为10.20 kJ·g-1.未经预处理的甲烷产出能量为26.01 kJ·g-1, 经过水热预处理的甲烷能量产出为27.88 kJ·g-1, 而经过臭氧预处理的甲烷能量产出为30.54 kJ·g-1.从能量平衡的角度上来看, 经过水热或臭氧预处理增加的甲烷能量输出(1.87 kJ·g-1以及4.53 kJ·g-1)无法弥补水热或臭氧预处理的能量消耗.但是经过预处理, 尤其是臭氧预处理后, 油脂降解速率加快, 达到产甲烷峰值速率的时间缩短, 提高了废弃油脂的发酵处理能力, 说明臭氧预处理对于废弃油脂的处理有很高的经济价值.
 |
| 图 4 3种油脂臭氧/水热预处理后厌氧发酵产甲烷过程总能量转化效率 Fig. 4 Energy conversion efficiency of pretreated waste lipids during anaerobic digestion |
由于餐厨垃圾以及其中的餐厨废油的成分和性质会由于来源地区, 收运时间地点等因素而改变, 文献中餐厨垃圾和餐厨废油的组成成分区别较大, 导致生物燃气和能量产率也有所差异.不过文献也表明, 一些预处理方式可以促进油脂降解, 促进厌氧发酵制生物甲烷.有学者采用脂肪酶或者生物处理法降解油脂, 能量转化效率为53.8%~61.9% (Meng et al., 2015; Liu et al., 2018).另外有研究表明, 臭氧预处理降解餐厨垃圾用于厌氧发酵能量转化效率约为58.8% (Javkhlan Ariunbaatar et al., 2014).本研究采用臭氧和水热预处理方式, 火锅油脂降解并厌氧发酵产甲烷的最高能量转化效率可达76.18%.在甲烷产率以及整体能量转化效率等参数方面已经达到或者超过使用酶或细菌进行预处理的油脂厌氧发酵甲烷产率及能量转化效率.
4 结论(Conclusions)本文以餐厨垃圾中典型油脂火锅油脂、烧烤油脂、川菜油脂作为发酵底物, 分别将臭氧预处理以及水热预处理的油脂用于厌氧发酵产甲烷以提高能量转化效率的实验研究.经过臭氧或水热预处理后, 火锅油脂厌氧发酵的甲烷产率分别达到了854.20 mL·g-1及779.72 mL·g-1, 整体能量转化效率分别由64.88%提升至76.18%和69.53%.3种油脂发酵产甲烷呈现了相似的规律, 且臭氧优于水热预处理.这说明油脂的臭氧及水热预处理是一个能够同时提高甲烷产率和能量转化效率的很有效的方法, 能够促进底物降解, 缓解油脂对于菌群的包埋.更多的研究应当着眼于降低水热及臭氧预处理能耗.
Adeva-Andany M M, Carneiro-Freire N, Seco-Filgueira M, et al. 2019. Mitochondrial beta-oxidation of saturated fatty acids in humans[J]. Mitochondrion, 46: 73-90. DOI:10.1016/j.mito.2018.02.009 |
Bong C P C, Lim L Y, Lee C T, et al. 2018. The characterisation and treatment of food waste for improvement of biogas production during anaerobic digestion: A review[J]. Journal of Cleaner Production, 172: 1545-1558. DOI:10.1016/j.jclepro.2017.10.199 |
Braguglia C M, Gallipoli A, Gianico A, et al. 2018. Anaerobic bioconversion of food waste into energy: A critical review[J]. Bioresource Technology, 248(Pt A): 37-56. |
Cheng J, Ding L, Lin R, et al. 2016a. Physicochemical characterization of typical municipal solid wastes for fermentative hydrogen and methane co-production[J]. Energy Conversion and Management, 117: 297-304. DOI:10.1016/j.enconman.2016.03.016 |
Cheng J, Su H, Zhou J, et al. 2011. Hydrogen production by mixed bacteria through dark and photo fermentation[J]. International Journal of Hydrogen Energy, 36(1): 450-457. DOI:10.1016/j.ijhydene.2010.10.007 |
Cheng J, Yue L, Ding L, et al. 2019a. Improving fermentative hydrogen and methane production from an algal bloom through hydrothermal/steam acid pretreatment[J]. International Journal of Hydrogen Energy, 44(12): 5812-5820. DOI:10.1016/j.ijhydene.2019.01.046 |
Cheng J, Yue L, Hua J, et al. 2019b. Hydrothermal heating with sulphuric acid contributes to improved fermentative hydrogen and methane co-generation from Dianchi Lake algal bloom[J]. Energy Conversion and Management, 192: 282-291. DOI:10.1016/j.enconman.2019.04.003 |
Cheng J, Yue L, Hua J, et al. 2020. Hydrothermal alkali pretreatment contributes to fermentative methane production of a typical lipid from food waste through co-production of hydrogen with methane[J]. Bioresource Technology, 306: 123164-123171. DOI:10.1016/j.biortech.2020.123164 |
De G G, Muntoni A, Polettini A, et al. 2017. Energy recovery from one- and two-stage anaerobic digestion of food waste[J]. Waste Management, 68: 595-602. DOI:10.1016/j.wasman.2017.06.013 |
Ding L, Cheng J, Lu H, et al. 2017a. Three-stage gaseous biofuel production combining dark hydrogen, photo hydrogen, and methane fermentation using wet Arthrospira platensis cultivated under high CO2 and sodium stress[J]. Energy Conversion and Management, 148: 394-404. DOI:10.1016/j.enconman.2017.05.079 |
Ding L, Cheng J, Qiao D, et al. 2017b. Investigating hydrothermal pretreatment of food waste for two-stage fermentative hydrogen and methane co-production[J]. Bioresource Technology, 241: 491-499. DOI:10.1016/j.biortech.2017.05.114 |
Li D, Yuan Z, Sun Y, et al. 2009. Hydrogen production characteristics of the organic fraction of municipal solid wastes by anaerobic mixed culture fermentation[J]. International Journal of Hydrogen Energy, 34(2): 812-820. DOI:10.1016/j.ijhydene.2008.11.031 |
Fisgativa H, Tremier A, Dabert P. 2016. Characterizing the variability of food waste quality: A need for efficient valorisation through anaerobic digestion[J]. Waste Management, 50: 264-274. DOI:10.1016/j.wasman.2016.01.041 |
Francioso O, Rodriguez-Estrada M T, Montecchio D, et al. 2010. Chemical characterization of municipal wastewater sludges produced by two-phase anaerobic digestion for biogas production[J]. Journal of Hazard Material, 175(1/3): 740-746. |
Jarunglumlert T, Prommuak C, Putmai N, et al. 2018. Scaling-up bio-hydrogen production from food waste: Feasibilities and challenges[J]. International Journal of Hydrogen Energy, 43(2): 634-648. DOI:10.1016/j.ijhydene.2017.10.013 |
Javkhlan A A P, Luigi F, Giovanni E, et al. 2014. Enhanced anaerobic digestion of food waste by thermal and ozonation pretreatment methods[J]. Journal of Environmental Management, 146: 142-149. DOI:10.1016/j.jenvman.2014.07.042 |
Kallscheuer N, Polen T, Bott M, et al. 2017. Reversal of beta-oxidative pathways for the microbial production of chemicals and polymer building blocks[J]. Metabolic Engineering, 42: 33-42. DOI:10.1016/j.ymben.2017.05.004 |
Li L, Peng X, Wang X, et al. 2018a. Anaerobic digestion of food waste: A review focusing on process stability[J]. Bioresource Technology, 248(Pt A): 20-28. |
Lin R, Cheng J, Murphy J D, et al. 2017. Unexpectedly low biohydrogen yields in co-fermentation of acid pretreated cassava residue and swine manure[J]. Energy Conversion and Management, 151: 553-561. DOI:10.1016/j.enconman.2017.09.006 |
Liu P, Ji J, Wu Q, et al. 2018. Klebsiella pneumoniae sp. LZU10 degrades oil in food waste and enhances methane production from co-digestion of food waste and straw[J]. International Biodeterioration & Biodegradation, 126: 28-36. |
Meng Y, Li S, Yuan H, et al. 2015. Evaluating biomethane production from anaerobic mono- and co-digestion of food waste and floatable oil (FO) skimmed from food waste[J]. Bioresource Technology, 185: 7-13. DOI:10.1016/j.biortech.2015.02.036 |
Xia A, Cheng J, Ding L, et al. 2015. Substrate consumption and hydrogen production via co-fermentation of monomers derived from carbohydrates and proteins in biomass wastes[J]. Applied Energy, 139: 9-16. DOI:10.1016/j.apenergy.2014.11.016 |
Xia A, Cheng J, Ding L, et al. 2014. Enhancement of energy production efficiency from mixed biomass of Chlorella pyrenoidosa and cassava starch through combined hydrogen fermentation and methanogenesis[J]. Applied Energy, 120: 23-30. DOI:10.1016/j.apenergy.2014.01.045 |
Xiao B, Qin Y, Wu J, et al. 2018. Comparison of single-stage and two-stage thermophilic anaerobic digestion of food waste: Performance, energy balance and reaction process[J]. Energy Conversion and Management, 156: 215-223. DOI:10.1016/j.enconman.2017.10.092 |
Yue L, Cheng J, Hua J, et al. 2020. Improving fermentative methane production of glycerol trioleate and food waste pretreated with ozone through two-stage dark hydrogen fermentation and anaerobic digestion[J]. Energy Conversion and Management, 203: 112225-112237. DOI:10.1016/j.enconman.2019.112225 |
Yun Y M, Lee M K, Im S W, et al. 2018. Biohydrogen production from food waste: Current status, limitations, and future perspectives[J]. Bioresource Technology, 248(Pt A): 79-87. |
Zhang W, Lang Q, Fang M, et al. 2017b. Combined effect of crude fat content and initial substrate concentration on batch anaerobic digestion characteristics of food waste[J]. Bioresource Technology, 232: 304-312. DOI:10.1016/j.biortech.2017.02.039 |