 2021, Vol. 41
2021, Vol. 41



2. 广东省微生物分析检测中心 广州 510070;
3. 华南应用微生物国家重点实验室 广州 510070;
4. 广东省菌种保藏与应用重点实验室 广州 510070
2. Guangdong Detection Center of Microbiology, Guangzhou 510070;
3. State Key Laboratory of Applied Microbiology Southern China, Guangzhou 510070;
4. Guangdong Provincial Key Laboratory of Microbial Culture Collection and Application, Guangzhou 510070
随着工农业发展和人类活动加剧, 水体重金属污染日益严重.引起水体污染的重金属主要包括镉、铬、镍、铅、汞、砷等生物毒性显著的元素.水体中重金属经农作物和饮用水进入人体并长期累积后, 严重威胁人体健康, 治理水体中重金属污染日益受到人们重视.传统重金属治理方法主要有物理法、化学法和生物法, 物理法主要包括离子交换和膜分离法, 处理条件受限且成本较高; 化学法包括沉淀法和电解法, 但容易对水体形成二次污染; 生物法一般选用高耐受重金属植物对土壤和水体中的重金属污染进行修复, 但处理周期较长.采用微生物吸附水体中重金属离子具有对环境毒害小(薛高尚等, 2012; Yu et al., 2015; 张冬雪等, 2017), 材料来源广泛, 成本低且可再生等优点而受到研究者关注(张泰然等, 2014; 李敏等, 2017).
酵母菌生长快、比表面积大、密度高, 营养需求简单、成本低(Hosseini et al., 2016; Camargo et al., 2018; Junior et al., 2019), 且其对重金属、pH、温度和渗透压耐受性强, 因而成为吸附重金属的首选微生物(Zhu et al., 2015; Ilyas et al., 2017; 李丽杰等, 2019; 孙禹, 2019).但目前大部分研究选用筛选的废弃酿酒酵母菌进行重金属吸附(孙禹, 2019), 关于毕赤酵母吸附重金属离子的研究还比较少, 主要聚焦于毕赤酵母吸附镉离子及其抗性机制研究(李春生, 2015).毕赤酵母主要被用于目的基因的异源表达(Jiang et al., 2016; Yan et al., 2019), 密度更高, 对营养需求简单, 便于大规模培养和成本控制, 因此毕赤酵母相对酿酒酵母在污水重金属离子处理方面有较好的应用前景.而且目前关于含特定基因的重组酵母菌对重金属离子吸附, 以及对酵母菌如何重复用于吸附重金属离子的研究报道较少.vgb基因编码透明颤菌血红蛋白, 该蛋白具有提高呼吸链传递速率提高溶解氧利用率的功能(Ma et al., 2014; Ye et al., 2019), 能促进菌体在贫氧条件下的生长和代谢, 同时提高代谢产物生成相关基因的表达水平(Kunkel et al., 2015; Ouyang et al., 2018), 因此vgb基因被广泛应用于提高目的基因表达水平和提升次级代谢产物产量(Jeong et al., 2015; Tang et al., 2016; Zhao et al., 2017).而污水中大多溶解氧有限, 因此, 含有vgb基因的重组酵母菌在污水处理方面有较好的应用前景.
本文选用野生毕赤酵母KM71和含有vgb基因的重组酵母KM71对重金属离子Ni2+、Cd2+和Pb2+进行吸附解吸试验, 研究其吸附解吸动力学特征及吸附剂用量、温度、pH值、混合金属离子的选择性吸附和解吸剂等因素对吸附率的影响, 并测定重金属离子胁迫下野生酵母菌和重组酵母菌的生长曲线.结果显示, 重组酵母菌比野生酵母菌具有更好的重金属吸附量和更强的重金属抗性, 采用不同解吸剂对吸附剂进行解吸时, 乙二胺四乙酸二钠(EDTA-2Na)对重金属离子的解吸效果相对较好.通过优化条件提升了重组酵母对重金属离子的吸附量, 将吸附重金属后的吸附剂用EDTA-2Na进行解吸-水洗-离心, 得到可重复利用的酵母并提升其吸附量.本研究为重组毕赤酵母作为重金属吸附剂及其重复利用提供了科学依据.
2 材料和方法(Materials and methods) 2.1 实验用菌株野生毕赤酵母KM71购买于武汉淼灵生物科技有限公司, 重组酵母菌为导入载体pPIC9K-vgb的KM71菌株(由本实验室构建).其中重组KM71菌株将在培养24 h后加入1%甲醇, 以诱导vgb基因的表达, 然后野生酵母和重组酵母菌均在培养48 h后收集菌体.培养液呈浑浊状, pH值为5.0左右, 摇匀后测定OD600值, 调整两种菌体的OD600值一致.试验用菌液的OD600值为10.0, 作为后续吸附实验的吸附剂(野生酵母Y-KM71, 重组酵母C-KM71).精密移取2 mL酵母菌液, 离心, 水洗后于75 ℃干燥, 得到2 mL菌液的干重为12 mg.
2.2 实验试剂和仪器HCl、NaOH、EDTA-2Na、硅藻土、活性炭、硝酸; pH计、多功能酶标仪用于测定生长曲线(Tecan, SPARK)、紫外分光光度计(T6, 北京普析通用)、离心机(ST16R, Thermo).
采用电感耦合等离子质谱仪(Agilent7800)测定重金属离子的浓度, 所有实验溶液在8000 r·min-1条件下离心3 min, 移取上清溶液用5%硝酸溶液稀释后测试浓度, 每个样品3个重复.
2.3 重金属溶液配制分别用氯化镉半(五水合物)(纯度98%, 麦克林); 氯化铅(纯度99.5%, 麦克林), 氯化镍(Ⅱ)(无水)(纯度98.0%, TCI)配制3个单元素的贮备溶液, 用贮备溶液经去离子水稀释分别得到单元素的金属溶液, 浓度分别为29.1、29.1、23.6 mg·L-1, 作为单元素吸附解吸实验溶液; 采用单元素贮备溶液经去离子水稀释得到二元金属混合溶液和三元金属混合溶液用于环境因素影响试验, 浓度约为30 mg·L-1; 配制不同浓度的三元金属混合溶液用于吸附等温线试验, 浓度为4~50 mg·L-1, 试验前测试溶液中金属离子实际浓度.
2.4 实验方法 2.4.1 不同浓度重金属离子对吸附剂的生长抑制影响在培养基中接种Y-KM71和C-KM71, 添加1 mg·L-1和5 mg·L-1的单一元素重金属溶液, 采用多功能酶标仪连续培养48 h测定酵母菌在不同浓度重金属胁迫下的生长曲线.
2.4.2 吸附解吸平衡时间及解吸剂选择精密移取3种单一元素的重金属溶液25 mL于50 mL塑料离心管中, 各加入2mL Y-KM71和C-KM71, 于25 ℃, 200 r·min-1振荡, 分别于15、30、60、120、180、240、300 min离心取样测试, 确定吸附平衡时间.分离上清液, 选择1%HCl, 0.05 mol·L-1 NaOH和0.05 mol·L-1 EDTA-2Na 3种解吸剂对吸附剂进行解吸, 确定解吸平衡时间和解吸率.
2.4.3 混合金属离子对吸附剂选择性吸附的影响精密移取二元金属混合溶液Ni2+-Cd2+、Pb2+-Cd2+、Ni2+-Pb2+和三元金属混合溶液Ni2+-Cd2+-Pb2+溶液25 mL于50 mL塑料离心管中, 各加入2 mL Y-KM71和C-KM71, 25 ℃, 200 r·min-1振荡120 min后测试浓度.
2.4.4 温度、pH值和不同吸附剂添加量对三元混合金属离子吸附性的影响精密移取三元混合金属溶液25 mL于50 mL塑料离心管中, 各加入2 mL Y-KM71和C-KM71, 于5、25和30 ℃静置120 min后测试浓度, 测试不同温度对吸附率影响.按相同步骤用1:1盐酸和1 mol·L-1NaOH调整溶液pH值为3、5、7和9, 各加入2 mL Y-KM71和C-KM71, 于25 ℃, 200 r·min-1振荡120 min后测试pH值对吸附率的影响.取等体积三元混合金属溶液, 分别添加0.50、1.0、2.0、5.0、10.0 mL的Y-KM71和C-KM71, 于25 ℃, 200 r·min-1振荡120 min后测试不同吸附剂添加量对吸附率和吸附量的影响.
2.4.5 不同浓度金属离子的吸附等温线精密移取不同浓度的吸附等温线金属溶液25 mL于50 mL塑料离心管中, 各加入2 mL Y-KM71和C-KM71, 于25 ℃, 200 r·min-1振荡120 min后取样测试, 用单元素贮备液配制的不同浓度梯度的三元混合金属溶液的实际浓度如表 1所示.
| 表 1 吸附等温线各金属离子浓度 Table 1 Concentration of metal ions in the adsorption isotherm |
根据两种酵母菌吸附重金属的结果, 选用0.01、0.02和0.03 g硅藻土和活性炭与2 mL C-KM71复合, 加入25 mL重金属溶液, 调节pH值为5.0、6.0、7.0, 吸附平衡后离心取样测试金属离子浓度, 分离上清液, 用0.05 mol·L-1 EDTA-2Na对吸附剂进行解吸2 h离心分离上清液, 用50 mL去离子水将吸附剂水洗-离心3遍, 加入25 mL重金属溶液吸附120 min后取样测试重复利用的KM71菌株对重金属离子的吸附率.
2.5 数据处理酵母菌对重金属离子的吸附率按式(1)计算.

|
(1) |
酵母菌对重金属离子的吸附量按式(2)计算.

|
(2) |
式中, C0为实验用重金属质量浓度(mg·L-1); V0为实验用重金属溶液体积(mL); C为吸附平衡后溶液中重金属质量浓度(mg·L-1); V为重金属溶液和添加酵母菌的体积和(mL); m为实验中添加的酵母菌重量(mg).
选择常用的Freundlich模型和Langmuir模型对吸附等温线数据进行拟合, Freundlich模型方程式为:
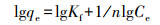
|
(3) |
Langmuir模型方程式为:
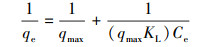
|
(4) |
式中, qe和Ce分别为吸附平衡时单位质量吸附剂吸附重金属离子的量和水相中金属离子浓度, Kf为Freundlich吸附系数, n为吸附回归常数, qmax为金属离子在单位质量吸附剂表面完全形成单层结构时的量, 代表吸附剂表面完全被金属离子覆盖时吸附剂的实际最大吸附能力, KL为吸附点位对重金属的结合力常数.
3 结果与讨论(Results and discussion) 3.1 重金属离子胁迫下对酵母菌生长的影响以重组载体pBEP43-vgb(本实验室保存)为模板, 扩增vgb基因, 酶切后插入至PPIC9K载体.菌液PCR筛选阳性克隆并测序验证.将pPIC9K-vgb载体(图 1a)用SalI进行线性化并进行回收, 然后在1.5 kV电压下电转至KM71感受态中.然后用组氨酸缺陷型的MD平板筛选阳性克隆.各菌落扩大培养, 提取总DNA, 扩增vgb基因并测序验证重组质粒PPIC9k-vgb的导入(图 1b).从而最终得到含有PPIC9K-vgb的重组毕赤酵母KM71菌株.将C-KM71和Y-KM71菌液进行超声破碎, 然后采用CO差光谱法对vgb基因所编码的透明颤菌血红蛋白(VHb)进行功能分析, 结果表明, C-KM71中可检测到419 nm处的特征吸收峰, 而Y-KM71则无法检测到相应的VHb特征吸收峰, 从而证实了VHb在重组C-KM71中的表达(图 1c).
 |
| 图 1 重组载体PPIC9K-vgb导入毕赤酵母KM71(a.PPIC9K-vgb重组载体图; b.PPIC9K-vgb导入KM71的菌液PCR验证; c. 重组KM71中VHb功能的验证) Fig. 1 The introduction of recombinant vector PPIC9k-vgb into Pichia pastoris KM71(a. the vector map of PPIC9k-vgb; b. the colony PCR verification of the introduction of PPIC9k-vgb; c.the verification of VHb function in C-KM71) |
根据生长曲线测定结果, 毕赤酵母KM71在5 h进入对数生长期, 到40 h进入平稳期. 1 mg·L-1的重金属离子Cd2+、Ni2+和Pb2+以及5 mg·L-1 Pb2+对野生酵母菌的生长没有影响,说明野生毕赤酵母KM71具有一定的重金属离子抗性,且Pb2+对KM71野生KM71菌株的生长抑制作用较小. 5 mg·L-1重金属离子Cd2+、Ni2+对重组毕赤酵母KM71(C-KM71)的生长几乎没有抑制作用, 而同等浓度重金属离子对野生毕赤酵母KM71(Y-KM71)的生长表现出一定的抑制作用.证实C-KM71对重金属离子具有更强的抗性.可能是因为重组毕赤酵母KM71中含有vgb基因, 能在周质空间和细胞膜表达透明颤菌血红蛋白, 根据Kalio的易化扩散假说和吴奕的末端电子受体假说, VHb与氧在周质空间和细胞膜的结合能提升重组毕赤酵母KM71的呼吸链传递效率和能量利用率, 同时也可能提升了金属硫结合蛋白等金属离子结合相关蛋白的表达水平, 因此, C-KM71对重金属离子的胁迫具有更好的抗性, 从而有利于重组毕赤酵母在高浓度重金属污染废水处理中的应用.
3.3 吸附剂对重金属离子的吸附特性 3.3.1 吸附剂对单一金属离子吸附解吸特性及解吸剂选择通过测定不同时间样品中重金属浓度来确定吸附解吸的平衡时间, 野生毕赤酵母和重组毕赤酵母对重金属的吸附在15 min内达到吸附平衡的60%; 在60 min能达到平衡时的80%, 在120 min后达到吸附平衡.这是因为酵母菌吸附重金属离子一般分为两个阶段, 第1阶段是细胞表面吸附阶段, 即胞外多聚物和细胞壁上的官能团(羧基、磷酰基、羟基、硫酸酯基和酰胺基)与金属离子发生配位络合, 该阶段快速、可逆且不依赖于能量代谢; 第2阶段是细胞表面的重金属离子在细胞表面相关酶的作用下转移至细胞内并参与细胞代谢的过程.吸附平衡时野生酵母Y-KM71对Ni2+、Cd2+和Pb2+的吸附率分别为18.80%、26.12%和46.59%, C-KM71对Ni2+、Cd2+和Pb2+的吸附率分别为22.80%、29.63%和52.04%(图 2).重组KM71对重金属的吸附效率高于野生KM71, 其原因为重组毕赤酵母KM71中的vgb基因在甲醇诱导下表达透明颤菌血红蛋白, 位于周质空间和细胞膜上的该蛋白可以提升重组KM71的能量传递效率和溶解氧利用率, 从而使C-KM71有更快的生长速率及更高的金属结合相关蛋白的表达水平.重组毕赤酵母C-KM71对Pb2+的吸附率显著高于Cd2+, 对Ni2+的吸附率最低.说明毕赤酵母对重金属的吸附具有选择性, 这主要与金属离子的离子半径和电负性有关, 相同价位的金属离子, 离子半径越大, 其水合半径越小, 越易被吸收, 吸附剂对Pb2+具有明显的选择性, 可能时由于Pb2+半径(0.132)较其他离子大, 且电负性(2.33)大于其他离子; 也可能由于铅离子对于KM71菌株的相对生物毒性较小, KM71对于铅离子的耐受性较强.为了便于后续对比实验, 因此选择添加2 mL
 |
| 图 2 C-KM71和Y-KM71对Ni2+、Cd2+和Pb2+的吸附动力学图 Fig. 2 Adsorption kinetics of Ni2+, Cd2+ and Pb2+ by C-KM71 and Y-KM71 |
吸附剂, 吸附平衡120 min.
采用1%HCl、0.05 mol·L-1 NaOH和0.05 mol·L-1 EDTA-2Na 3种解吸剂对重金属离子Pb2+、Cd2+、Ni2+进行解吸, 在60 min达到解吸平衡.结果如表 2所示, 0.05 mol·L-1 EDTA-2Na对Ni2+和Pb2+的解吸结果相对更好, 1% HCl对Cd2+的解吸结果相对更好.这几种解吸剂不能将金属离子从毕赤酵母KM71中完全解吸, 同时也证实了毕赤酵母KM71对重金属离子良好的吸附性能.根据解吸结果, 后续解吸实验的解吸剂选用0.05 mol·L-1 EDTA-2Na, 解吸平衡时间为1 h.
| 表 2 吸附剂对金属离子的解吸结果 Table 2 Desorption of metal ions by adsorbents |
混合体系的吸附动力学用qe′/qe表示, qe′/qe代表一种元素在混合体系中与单一元素存在时吸附量的比值.qe′/qe﹥1表示两种离子之间具有协同作用, qe′/qe < 1表示两种离子间存在着拮抗作用, qe′/qe=1表示两种离子间无作用.二元体系Ni-Cd下, Cd的qe′/qe﹥1, 分别为1.95和1.89, Ni2+的存在能促进Cd2+的吸附, 其余的二元体系中qe′/qe < 1, 特别是Cd2+-Pb2+体系中, 两种金属离子的qe′/qe非常低, 暗示这两种金属离子间存在较强的拮抗作用, 可能由于其存在相同的吸附位点和基团.在三元体系中, 其qe′/qe均小于1, 吸附剂对3种离子的吸附能力顺序是:Pb2+﹥Cd2+﹥Ni2+.说明毕赤酵母KM71菌株吸附Pb2+、Cd2+、Ni2+这3种重金属离子时, 这3种重金属离子存在较强的拮抗作用.
3.3.3 温度和pH值金属离子吸附性的影响不同温度条件下, 野生和重组KM71菌株对重金属离子的吸附率差异不明显, 在5、25和30 ℃试验条件下, Y-KM71和C-KM71对几种重金属离子的吸附率变化菌小于5%.说明Y-KM71和C-KM71具有宽泛的温度适应性, 从而有利于其在不同水体环境中的重金属离子污染治理.在pH值为3.0、5.0、7.0和9.0时, 野生和重组KM71菌株对重金属离子的吸附率先升高后降低, 总体差异较小(图 3).在低pH条件下, 重金属的吸附率相对偏低, 可能是由于溶液中大量H+与重金属离子争夺吸附位点, 同时阻碍活性基团的解离; KM71菌株对重金属离子的吸附率随着pH值升高而升高, 在pH 5.0~7.0时, 吸附率达到最高, 这可能是溶液中H+急剧减少, 降低了与金属离子对吸附剂吸附位点的竞争, 带负电荷的吸附剂与金属离子的静电作用加强, 故有利于对金属离子的吸附.在pH 9.0条件下, 吸附剂对重金属离子的吸附率相对有所下降, 这可能与酵母菌自身的生长代谢相关.酵母菌的生长pH为5.0左右, 当pH为9.0时, 过碱的环境不利于酵母菌的生长及次级代谢, 其吸附重金属离子的能力有所降低.且在碱性条件下而金属离子易形成不溶解的氧化物、氢氧化物等沉淀, 覆盖于KM71酵母菌株表面, 也不利于金属离子的吸附.酵母菌具有较宽的pH吸附值, 同等条件下, 通过添加NaOH适当调节溶液pH值至中性可提高KM71菌株对重金属离子的吸附率.
 |
| 图 3 pH值对C-KM71和Y-KM71吸附Ni2+、Cd2+和Pb2+的影响 Fig. 3 Effect of pH value on the adsorption of Ni2+, Cd2+ and Pb2+ by C-KM71 and Y-KM71 |
当添加不同剂量的野生和重组KM71菌株后, 吸附平衡后的溶液中金属离子的浓度随吸附剂的增加而减少, 且KM71菌株对重金属离子的吸附率随着菌体添加量的增加而提升.但单位质量的菌体对重金属离子的吸附量随菌液添加量的增大而明显减少(图 4).因为当KM71菌株对金属离子吸附总量一致时, 当添加吸附剂的量越多时, 体系总体积增大, 而且菌体添加量过多时KM71菌株对重金属离子的吸附存在一定的竞争效应, 单位体积菌体吸附的离子量就少.李丽杰等(2020)采用异常威客汉姆酵母QF-1-1吸附Pb2+, 在Pb2+溶液初始pH 5.5、Pb2+溶液浓度为100 mg·L-1、菌体质量浓度为11 g·L-1条件下, 吸附率为97.89%, 吸附量为7.29 mg·g-1, 该吸附量远低于本试验中C-KM71对Pb2+的吸附量.实验添加的菌量约为0.48 g·L-1.其对重金属离子的吸单位质量菌体吸附量也会随着菌体添加量的增加而降低.
 |
| 图 4 吸附剂添加量对C-KM71和Y-KM71吸附Ni2+、Cd2+和Pb2+的影响 Fig. 4 Effect of adsorbent addition on the adsorption of Ni2+, Cd2+ and Pb2+ by C-KM71 and Y-KM71 |
KM71菌株对2.4.5节中不同离子浓度的重金属吸附特征选择常用的Freundlich模型和Langmuir模型对等温线数据进行拟合, 得到等温吸附模型的相关参数如表 3所示, Langmuir模型用于描述吸附质在含有有限的同类吸附点位的表面均一的吸附剂上发生的单分子层吸附, 拟合的线性相关系数的平均值为0.9520, 但是相关参数未能反映真实的吸附情况, 特别是重组菌的qmax出现负值, 说明吸附过程不是简单的单分子层吸附, 存在复杂的胞内吸附.采用Freundlich模型的线性相关系数的平均值为0.9448, 线性拟合结果的回归常数n均大于1, 1/n的值在0.7271~0.8906之间, 表明在等温吸附的初始阶段, 酵母对金属离子有较强的亲和力, 随着酵母上吸附位点逐渐被占领, 金属离子的吸附位点越来越少, 重组毕赤酵母C-KM71对Cd2+和Pb2+的吸附强度明显高于野生酵母菌, Freundlich模型的吸附等温线图如图 5所示, 单位质量吸附剂对Ni2+、Cd2+和Pb2+的吸附量均随着溶液中离子浓度的增大而升高, 到一定浓度时达到平衡, 且重组C-KM71菌株对金属离子的吸附力高于野生Y-KM71菌株.
| 表 3 C-KM71吸附重金属离子的模型参数 Table 3 Model parameters for adsorption of heavy metal ions by C-KM71 |
 |
| 图 5 C-KM71和Y-KM71对Ni2+、Cd2+和Pb2+的吸附等温线 Fig. 5 Adsorption isotherms of Ni2+, Cd2+ and Pb2+ on C-KM71 and Y-KM71 |
前期实验结果表明, C-KM71对重金属离子的吸附力优于Y-KM71, 选用硅藻土和活性炭作为C-KM71的复合剂, 可为C-KM71提供载体.载体表面的基团也可以与重金属离子结合, 形成共价键和离子键.载体空白对照、载体与C-KM71复合对重金属离子的吸附结果如表 4所示, 活性炭对重金属离子Ni2+、Cd2+和Pb2+的吸附能力优于硅藻土.选用0.03 g活性炭与C-KM71复合, 复合活性炭-C-KM71对重金属离子的吸附率大于两者分别对重金属离子吸附率之和.这可能是因为复合剂为微生物生长提供了载体, 且由于本身具有的多孔结构和富含活性基团, 对重金属离子具有一定的富集作用, 因此对于C-KM71的吸附重金属性能有一定的提升.
| 表 4 C-KM71吸附重金属离子的条件优化结果 Table 4 Optimization of conditions for adsorption of heavy metal ions by C-KM71 |
调整C-KM71-活性炭吸附重金属离子的pH值为5.0、6.0和7.0, 吸附剂经离心分离后, 采用EDTA-2Na进行解吸, 水洗3次得到可重复利用的吸附剂.C-KM71菌株对Ni2+、Cd2+和Pb2+二次吸附的结果中, pH值对其吸附率影响不大, 但相对比未经过水洗处理的C-KM71, 其对重金属离子的吸附能力得到了较大幅度的提升, 结果如表 4所示.水洗C-KM71菌株对Ni2+、Cd2+和Pb2+的最佳吸附pH值分别为6.0、6.0、5.0, 吸附率分别为35.20%、50.08%、69.84%, 相比未经处理的C-KM71, 其对重金属离子的吸附率分别提高了15.14%、21.04%、18.47%, C-KM71对Ni2+、Cd2+和Pb2+吸附量为21.27、25.08、39.87 mg·g-1, 相比水洗前分别提高了11.26、13.43、22.90 mg·g-1.该结果表明水洗能提升毕赤酵母C-KM71对重金属离子的吸附性能.主要可能是因为通过水洗能暴露出更多与吸附相关的官能团.代淑娟等(2008)采用扫描电镜与红外光谱等分析手段, 研究水洗废啤酒酵母吸附电镀水中的Cd2+, 结果表明对酵母水洗预处理后, 其对Cd2+吸附率由89.85%提高到96.18%, 其原因可能是水洗预处理去除了酵母中掩盖吸附位点的可溶相, 进而暴露出更多的吸附位点, 从而提高了酵母与金属离子发生配合作用的几率.
4 结论(Conclusions)vgb基因导入毕赤酵母KM71能提升其吸附重金属离子的能力, 因此非常适用于低溶氧污水中重金属的处理.在重金属离子吸附试验中, 重组毕赤酵母C-KM71的吸附性能优于野生毕赤酵母Y-KM71, 且重组毕赤酵母C-KM71相比Y-KM71具有更好的重金属抗性.通过条件优化能提高毕赤酵母KM71对重金属离子Ni2+、Cd2+和Pb2+的吸附率和吸附量, 重组毕赤酵母C-KM71与0.3 mg活性炭复合达到吸附平衡时, 其对Ni2+、Cd2+和Pb2+的最佳吸附pH值分别为6.0、6.0、5.0, 其对Ni2+、Cd2+和Pb2+相应吸附率分别为35.20%、50.08%、69.84%, 其对Ni2+、Cd2+和Pb2+的吸附量分别为21.27、25.08、39.87 mg·g-1.本论文提供了一种对重金属离子Ni2+、Cd2+和Pb2+高抗性及高吸附性能的含vgb的毕赤酵母KM71, 并研究了其对不同重金属离子的吸附解吸特性及动力学特征, 为毕赤酵母在重金属污染水体治理中的应用提供了可靠的理论依据.
Camargo F P, Prado P F D, Tonello P S, et al. 2018. Bioleaching of toxic metals from sewage sludge by co-inoculation of Acidithiobacillus and the biosurfactant-producing yeast Meyerozyma guilliermondii[J]. Journal of Environmental Management, 211(APR.1): 28-35. |
代淑娟, 高太, 王玉娟, 等. 2008. 共存离子对水洗废啤酒酵母吸附水相中Cd2+的影响[J]. 有色矿冶, 24(3): 79. DOI:10.3969/j.issn.1007-967X.2008.03.030 |
Hosseini M, Keshtkar A R, Moosavian M A. 2016. Electrospun chitosan/baker's yeast nanofibre adsorbent: preparation, characterization and application in heavy metal adsorption[J]. Bulletin of Materials Science, 39(4): 1091-1100. DOI:10.1007/s12034-016-1260-5 |
Ilyas S, Rehman A, Ilyas Q. 2017. Heavy metals induced oxidative stress in multi-metal tolerant yeast, Candida sp. PS33 and its capability to uptake heavy metals from wastewater[J]. Pakistan Journal of Zoology, 49(3): 769-775. DOI:10.17582/journal.pjz/2017.49.3.769.775 |
Jeong J W, Park K M, Chung M, et al. 2015. Influence of Vitreoscilla hemoglobin gene expression on 2, 3-butanediol production in Klebsiella oxytoca[J]. Biotechnology and Bioprocess Engineering, 20(1): 10-17. DOI:10.1007/s12257-014-0642-z |
Jiang C, Song J, Cong H, et al. 2016. Expression and characterization of a novel antifungal Exo-β-1, 3-glucanase from Chaetomium cupreum[J]. Applied Biochemistry & Biotechnology, 182(1): 1-15. |
Ji Q L, Li D, Li X Q, et al. 2015. Enhancement of Gellan Production in Sphingomonaspaucimobilis JLJ by Heterogeneous Expression of Vitreoscilla Hemoglobin[J]. Lecture Notes in Electrical Engineering, 332: 427-438. |
Junior R B D R, Meira H M, Almeida D G, et al. 2019. Application of a low-cost biosurfactant in heavy metal remediation processes[J]. Biodegradation, 30: 215-233. DOI:10.1007/s10532-018-9833-1 |
Kunkel S A, Pagilla K R, Stark B C. 2015. Engineering ofNitrosomonas europaeato expressVitreoscillahemoglobin enhances oxygen uptake and conversion of ammonia to nitrite[J]. Amb Express, 5(1): 43. DOI:10.1186/s13568-015-0135-2 |
李春生. 2015. 库德毕赤酵母重金属积累特性及高盐/低pH下镉抗性提高机理研究[D]. 青岛: 中国海洋大学
|
李丽杰, 贺敏, 贺银凤, 等. 2019. 酵母菌对重金属的吸附与抗性和解毒重金属的胞内分子机制研究进展[J]. 食品科学, 40(3): 330-337. |
李丽杰, 贺银凤. 2020. 高吸附Pb2+的异常威客汉姆酵母QF-1-1吸附Pb2+特性及机理[J]. 食品科学, 41(10): 152-158. |
李敏, 滕泽栋, 朱静, 等. 2017. 解磷微生物修复土壤重金属污染研究进展[J]. 生态学报, 38(10): 3393-3402. |
Ma R J, Lin X Z. 2014. Vitreoscilla hemoglobin gene (vgb) improves lutein production in Chlorella vulgaris[J]. Chinese Journal of Oceanology and Limnology, 32(2): 390-396. DOI:10.1007/s00343-014-3142-2 |
Ouyang P, Wang H, Hajnal I, et al. 2018. Increasing oxygen availability for improving poly(3-hydroxybutyrate) production by Halomonas[J]. Metabolic Engineering, 45: 20-31. DOI:10.1016/j.ymben.2017.11.006 |
孙禹. 2019. 高吸附Pb2+酵母菌的益生特性、吸附特性及吸附机理的研究[D]. 内蒙古: 内蒙古农业大学
|
Tang B, Feng X H, Zhang D, et al. 2016. Expression of Vitreoscilla hemoglobin gene to enhance poly-γ-glutamic acid biosynthesis in Bacillus subtilis NX-2[J]. Chinese Journal of Bioprocess Engineering, 2: 1-6. |
薛高尚, 胡丽娟, 田云, 等. 2012. 微生物修复技术在重金属污染治理中的研究进展[J]. 中国农学通报, 28(11): 266-271. |
Yan G Y, Yu L, He J L, et al. 2019. High efficiency induction and expression of oyster metallothionein recombinant Pichia pastoris strain[J]. Biotechnology & Business, 5: 64-70. |
Ye W, Zhang W M, Chen Y C, et al. 2016. A new approach for improving epothilone B yield in Sorangium cellulosumby the introduction ofvgbepoFgenes[J]. Journal of Industrial Microbiology & Biotechnology, 43(5): 641-650. DOI:10.1007%2Fs10295-016-1735-9 |
Yu Z Q. 2015. Advances in microbial remediation on theapplication of heavy metal pollution inagricultural water resources[J]. Agricultural Science & Technology, 16(12): 2824-2828. |
Zhao T X, Li M, Zheng X, et al. 2017. Improved production of trans-4-hydroxy-l-proline by chromosomal integration of the Vitreoscilla hemoglobin gene into recombinant Escherichia coli with expression of proline-4-hydroxylase[J]. Journal of Bioscience & Bioengineering, 123: 109-115. |
张冬雪, 丰来, 罗志威, 等. 2017. 土壤重金属污染的微生物修复研究进展[J]. 江西农业学报, 29(8): 62-67. |
张泰然, 王维. 2014. 铅污染及其微生物修复技术的研究进展[J]. 微生物前沿, 3: 1-7. |