 2021, Vol. 41
2021, Vol. 41



2. 哈尔滨工业大学环境学院, 哈尔滨 150090
2. School of Environment, Harbin Institute of Technology, Harbin 150090
随着经济的高速发展, 能源短缺问题日益突出, 可再生能源的生产迫在眉睫.在诸多的新型替代能源中, 氢气因具有热值高、无污染、可再生等优点而被认为是最具吸引力的石油替代能源(Yang et al., 2019).近年来, 由于生物制氢可以利用有机废水、农产品加工废弃物、食品垃圾等廉价原料通过微生物的作用发酵产氢而受到越来越多研究者的青睐(Arimi et al., 2015; Fuess et al., 2019).然而, 生物制氢效率低是限制其工业化生产的主要因素之一.因此, 强化厌氧污泥发酵产氢效能对于提高有机废弃物生物制氢效能具有重要意义.
厌氧污泥产氢效能的提高主要从两方面着手, 一是抑制耗氢菌的活性, 二是提高产氢菌的产氢能力.产甲烷菌等耗氢菌的存在会显著降低厌氧污泥的发酵产氢效能.因此在厌氧发酵制氢的过程中需要抑制耗氢微生物的活性.常见的产甲烷抑制剂包括2-溴乙酸磺酸钠(BES)、氯仿和替硝唑等(王晋等, 2011; 金鑫等, 2019).近年来, 氧化还原介体(ROMs)被广泛用于强化偶氮染料、多氯联苯等污染物的厌氧降解(李海波等, 2012; Zhang et al., 2017; 班巧英等, 2019).前期研究发现, 腐殖酸作为ROMs能够显著提高厌氧污泥的发酵产氢能力.为了考察腐殖酸和产甲烷抑制剂联合作用对厌氧污泥发酵产氢效能的影响, 本研究通过间歇试验考察了不同产甲烷抑制剂(氯仿、BES、替硝唑)和腐殖酸联合作用对厌氧污泥发酵葡萄糖产氢的强化效果, 并通过高通量测序揭示了产甲烷抑制剂和腐殖酸联合作用对产酸发酵相关微生物菌群的影响.
2 材料与方法(Materials and methods) 2.1 接种污泥和试验废水接种污泥取自太原市某城市污水处理厂的缺氧池, 将污泥混合液置于烧杯中静置6 h, 弃上清液后用于试验.污泥浓度为2.4 g·L-1.本研究中所用有机废水为人工合成废水, 以5000 mg·L-1葡萄糖作为唯一碳源, 并添加微生物生长繁殖所需要的其他营养元素(班巧英等, 2020).
2.2 腐殖酸和产甲烷抑制强化污泥产氢试验本研究腐殖酸的浓度设定为80 mg·L-1, 选用的产甲烷抑制剂为0.02%氯仿、0.04%氯仿、BES(10 mmol·L-1)和替硝唑(1 g·L-1).共设置10组试验:①空白对照、②腐殖酸、③0.02%氯仿、④0.02%氯仿+腐殖酸、⑤0.04%氯仿、⑥0.04%氯仿+腐殖酸、⑦BES、⑧BES+腐殖酸、⑨替硝唑、⑩替硝唑+腐殖酸.试验采用间歇培养方式进行, 实验容器为300 mL厌氧瓶.每个厌氧瓶中加入10 mL接种污泥和10 mL试验废水, 然后按照试验设计向厌氧瓶中加相应试剂, 最后加入适量蒸馏水使液相总体积达到100 mL.将培养液pH值用1 mol·L-1 HCl调至6.5.连续通入3 min N2后立即密封.将所有厌氧瓶置于35 ℃、120 r·min-1条件下恒温震荡培养.每组试验设置3个平行样, 数据分析取其平均值.每12 h取一次样测定其气体组成, 发酵结束时测定VFAs浓度.
2.3 Illumina MiSeq测序称取0.15 g污泥(湿重), 采用DNA提取试剂盒(E.Z.N.ATMMag-BindSoil DNA Kit, OmegaBio-tek, Inc., USA)提取污泥总DNA.以样品DNA为模板, 用通用引物27F:5′-AGAGTTTGATCMTGGCTCAG-3′, 515R:5′- CACGTATTACCGCGGCTGCTGGCAC-3′对细菌的16S rRNA基因V1~V3区进行PCR扩增.构建Illumina平台文库并测序.使用Mothur软件(version v.1.30.1)对测序获得的原始序列进行质控筛选和过滤, 按照97%相似性进行OTU(Operational Taxonomic Unit)聚类分析, 在聚类过程中去除嵌合体, 得到各OTU的代表序列, 在Silva数据库(http://www.arb-silva.de)进行比对、物种注释, 最后在门水平和属水平进行群落结构分析.
2.4 分析项目及方法生物量(挥发性悬浮固体总量MLVSS)采用标准方法测定(国家环保局, 2012), 产气量通过10~50 mL的玻璃注射器排气计量.发酵气体的组分和挥发酸浓度分别采用山东滕州瑞普分析仪器有限公司的RP-6800A型(TCD检测器)和RP-6890型(FID检测器)气相色谱测定.累计氢气产量参照Owen法进行计算(张立国等, 2016).
2.5 动力学分析产氢动力学分析参照Gompertz模型(1).
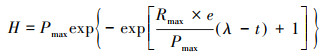
|
(1) |
式中, H为反应t(h)累计氢气产量(mL), Pmax为最大产氢量(mL), Rm ax为最大产氢速率(mL·h-1), λ为延迟时间(h).将累计氢气产量和相应反应时间t代入公式, 用统计软件Origin 9.0计算出Pmax、Rmax、λ(张立国等, 2016).
2.6 统计分析不同样品间差异显著性分析采用SPSS 22.0.0.0, Tukey HSD模型分析.
3 结果与讨论(Results and discussion) 3.1 累积氢气产量ROMs作为电子传递体, 可以提高电子从初级电子供体传递到最终电子受体的速度, 从而提高氧化还原反应速率(班巧英等, 2019).前期的研究发现, 腐殖酸能够显著提高厌氧污泥的产氢能力.而适当的产甲烷抑制剂的选择也会对厌氧污泥的产氢效能产生影响.因此, 本研究考察了腐殖酸与不同产甲烷菌抑制剂联合作用对厌氧污泥发酵产氢能力的影响.
如图 1所示, 在接种培养至第24 h时, 对照和腐殖酸发酵系统的累计产氢量达到最大值, 分别为10.6 mL和10.1 mL, 随后氢气产量迅速减少, 表明对照和腐殖酸发酵系统中存在耗氢菌, 且耗氢菌的活性高于产氢菌, 因而出现了氢气迅速减少的现象.从图 2可以看出, 这两个发酵系统中的甲烷含量随着培养时间的延续持续增加, 表明产甲烷菌为该发酵系统的主要耗氢菌.当发酵系统加入产甲烷抑制剂0.02%氯仿、0.04%氯仿和BES后, 产甲烷菌的活性受到抑制, 在整个培养过程均未检测到甲烷.经过120 h的培养后, 0.02%氯仿、0.04%氯仿和BES发酵系统的累计产氢量分别达到了15.7、13.2和9.4 mL.当发酵系统中同时加入腐殖酸和0.02%氯仿、0.04%氯仿或BES时, 厌氧污泥的产氢效能均高于单独添加产甲烷抑制剂的发酵系统.其中, 0.02%氯仿+腐殖酸发酵系统产氢效果最佳, 其累计氢气产量为17.4 mL, 显著高于其他发酵系统(p < 0.01).由此可见, 腐殖酸和产甲烷抑制剂的联合作用可以显著提高厌氧污泥的发酵产氢效能.腐殖酸强化厌氧污泥发酵产氢效能主要是由于它们能够降低葡萄糖氧化还原反应的活化能、提高产氢微生物的能量利用效率(班巧英等, 2019).然而, 当替硝唑作为产甲烷抑制剂, 两组试验(替硝唑和替硝唑+腐殖酸)均存在耗氢现象, 但是在整个发酵过程中并没有检测到甲烷(图 1).这可能是由于发酵系统中存在一些其他的耗氢菌, 如硫酸盐还原菌、同型产乙酸菌的等(Parameswaran et al., 2012; Rodrigues et al., 2020).
 |
| 图 1 不同产甲烷抑制剂和腐殖酸对累积产氢量的影响 Fig. 1 Effects of different methanogenesis inhibitors and humic acid on cumulative hydrogen production |
 |
| 图 2 对照和腐殖酸系统中甲烷含量变化曲线 Fig. 2 The change of methane content in the control and humic acid systems |
为了解各发酵系统的动力学特征, 采用改进的Gompertz模型(式(1))对累计氢气产量曲线进行了非线性拟合(表 1).如表 1所示, 除耗氢发酵系统外, 其余发酵系统的拟合系数(R2)均大于0.99, 表明该模型用来描述累计产氢量的进程是合理的.由表 1的拟合结果可以发现, 0.02%氯仿、0.04%氯仿和BES试验组的最大产氢量(Pmax)分别为16.1、13.8和12.2 mL.当系统中加入腐殖酸后Pmax比单独加产甲烷抑制剂高出8.0%~12.3%.其中0.02%氯仿+腐殖酸发酵系统的Pmax最高, 为17.7 mL.从最大产氢速率(Rmax)上看, 0.02%氯仿和0.02%氯仿+腐殖酸发酵系统具有相同的Rmax, 显著高于其他发酵系统(p < 0.05).另外, 拟合结果也表明, 腐殖酸的加入可以缩短迟滞期(λ).由此可见, 0.02%氯仿+腐殖酸组合不但可以抑制产甲烷菌的耗氢作用, 同时能够显著强化厌氧活性污泥的产氢能力.
| 表 1 产氢动力学分析 Table 1 Kinetic parameters for hydrogen production |
有机物厌氧发酵产氢过程通常伴随着挥发酸的产生(Abubackar et al., 2019).如图 3所示, 对照和腐殖酸系统中的液相末端产物以乙醇、乙酸和丙酸为主, 它们的含量分别为73.0/66.8、86.2/109.0和72.3/82.3 mg·L-1.类似地, 当0.02%氯仿和0.04%氯仿作为产甲烷抑制剂时, 乙醇、乙酸和丙酸仍然为主要液相末端产物, 但它们的浓度显著高于对照发酵系统(p < 0.01), 表明产甲烷抑制剂的添加可以提高产酸发酵菌群的活性.当在发酵系统中同时加入0.02%或0.04%氯仿和腐殖酸时, 丁酸含量分别达到了131.6和102.3 mg·L-1, 成为主要的液相发酵产物.相反, 这两个发酵系统中丙酸的含量显著减少(p < 0.01), 占总挥发酸的15%左右, 表明腐殖酸可以促进丁酸发酵菌的生长.然而, 当BES和替硝唑作为产甲烷抑制剂时, 无论是否加入腐殖酸, 乙酸和丙酸始终为主要的液相末端产物.此外, 0.02%氯仿+腐殖酸发酵系统的挥发酸总量也高于其他发酵系统(p < 0.01), 表明0.02%氯仿和腐殖酸能够提高产酸发酵菌群的活性, 从而提高累计产氢量(图 1).
 |
| 图 3 产甲烷抑制剂和腐殖酸对发酵液相末端产物的影响 (图中相同字母表示样品间挥发酸总量差异不显著, 不同字母代表样品间挥发酸总量差异显著) Fig. 3 Effect of methanogenesis inhibitors and humic acid on liquid fermentation end product |
厌氧活性污泥的发酵产氢效能与其群落结构密切相关(Qin et al., 2019).为了解腐殖酸与产甲烷抑制剂联合作用对葡萄糖发酵产氢气过程中微生物群落结构的影响, 本研究采用Illumina MiSeq测序技术对对照、0.02%氯仿+腐殖酸、0.04%氯仿+腐殖酸发酵系统中厌氧污泥进行了群落结构解析.如表 2所示, 经过筛选、剔除不合格序列后, 3个样品包含的有效序列数目分别为5.1×104、5.7×104、5.5×104.基于97%相似性将这些序列进行OTUs聚类分析, 在对照、0.02%氯仿+腐殖酸、0.04%氯仿+腐殖酸污泥样品中分别获得7.1×103、7.4×103、8.0×103个OTUs.从OTUs数目上看, 多样性由高到低依次为:0.04%氯仿+腐殖酸>0.02%氯仿+腐殖酸>对照.然而Simpson和Shannon多样性指数表明, 微生物群落多样性由大到小依次为0.04%氯仿+腐殖酸>对照>0.02%氯仿+腐殖酸.类似地, 0.04%氯仿+腐殖酸的丰富度指数(Chao1和Ace)显著高于对照和0.02%氯仿+腐殖酸.尽管Chao1和Ace估计的OTUs数目高于实际检测值, 然而文库覆盖率(Coverage)达到了89%以上, 表明样品中主要微生物已被检出.
| 表 2 微生物群落多样性和丰富度分析 Table 2 Analysis of diversity and richness of microbial commnity |
为揭示对照、0.02%氯仿+腐殖酸、0.04%氯仿+腐殖酸发酵系统中微生物群落结构的相似性和差异性, 对以上3个样品进行了主成分分析, 并通过Venn图揭示了OTUs分布特征(图 4).如图 4a所示, PC1和PC2的贡献率分别为50.9%和49.1%.PCA分析结果表明, 对照、0.02%氯仿+腐殖酸、0.04%氯仿+腐殖酸发酵系统中微生物群落组成具有显著差异.Venn图进一步揭示了3个样品中共有和独有的OTUs数目.由图 4b可知, 1150个OTUs存在于所有样品中, 仅占样品OTUs总数的6.1%.而对照、0.02%氯仿+腐殖酸、0.04%氯仿+腐殖酸样品中独有的OTUs数目分别为5165、5410和6040, 占21.2%~31.8%.
 |
| 图 4 不同发酵条件下主成分分析(a)和Venn图(b) Fig. 4 PCA analysis and Venn diagram at different fermentation conditions |
如图 5a所示, 在门水平上, 未分类的序列为9.27%~9.95%, 而已鉴定的序列在系统发育上涉及11个门.其中, 变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)、拟杆菌门(Bacteroidetes)、厚壁菌门(Firmicutes)和放线菌门(Actinobacteria) 为中温厌氧反应器中常见的水解发酵菌群(Maspolim etal., 2015;Antwi et al., 2017).与已有研究结果相似, 本研究发现变形菌门和拟杆菌门为所有样品的优势细菌门, 它们的相对丰度分别为47.3%~48.8%和11.1%~11.7%.在对照中, 厚壁菌门和放线菌门的相对丰度分别为3.9%和3.7%, 当加入腐殖酸和氯仿后, 这两个门的相对丰度显著增加, 比对照提高了18.9%~67.6%.相反, 硝化螺旋菌门(Nitrospirae)的相对丰度在对照中较高, 而在0.02%氯仿+腐殖酸和0.04%氯仿+腐殖酸中的相对丰度较低.其余真细菌门的相对丰度在各样品之间没有显著差异.除了真细菌外, 所有样品中均检测到少量广古菌门(Euryarchaeota)的微生物.研究表明, 已鉴定的产甲烷菌多数来自广古菌门.在对照中, 它的相对丰度为1.1%, 当加入氯仿后, 相对丰度显著降低, 表明氯仿可以有效抑制产甲烷菌的活性.
 |
| 图 5 不同发酵条件下细菌群落在门(a)和属(b) 水平的相对丰度 Fig. 5 Relative abundance of bacterial community at phylum and genus levels under different fermentation conditions |
从图 5b可以看出, 在属水平上未分类和其他序列的相对丰度高达51.5%~52.4%和24.5%~25.4%.由此可见, 这些未知功能的微生物种属在葡萄糖发酵产氢过程中起着重要作用.除了未知细菌属外, 高通量测序共检测到13个菌属.在对照组中, Ottowia(4.2%)、Ignavibacterium(2.8%)、Nitrospira(2.4%)、Saccharibacteria_ genera_incertae_sedis(2.2%)和Terrimonas(2.3%)为主要菌属.Ottowia可以利用小分子有机酸(如延胡索酸、乳酸)为碳源(Spring et al., 2004). Ignavibacterium、Saccharibacteria_genera_incertae_sedis和Terrimonas能够降解一些碳水化合物、大分子有机物等(Liu et al., 2012).Nitrospira在低溶解氧的条件下可产生硝化作用而消耗氢气(Lin et al., 2020).当加入0.02%氯仿和腐殖酸后, Ottowia和Saccharibacteria_genera_incertae_sedis的相对丰度有所增加, 而当加入0.04%氯仿和腐殖酸后他们的数量低于对照组, 表明较高浓度的氯仿对产酸发酵菌群的生长具有一定的抑制性.相反, 当加入氯仿后, Nitrospira的数量减少为0, 说明氯仿可以有效抑制这类耗氢菌的生长.除此之外, 氯仿和腐殖酸的加入还促进了Ilumatobacter和Ferruginibacter的生长.另外, 在对照中还检测到一些嗜氢产甲烷菌和嗜乙酸产甲烷菌, 它们的相对丰度分别为0.8%和0.4%, 当加入氯仿后, 显著减少.
4 结论(Conclusions)1) 0.02%氯仿、0.04%氯仿、BES和腐殖酸的联合作用均能提高厌氧污泥的发酵产氢能力, 其中0.02%氯仿和腐殖酸组合最佳, 其累计氢气产量达到了17.4 mL, 比对照提高了13.6倍, 且Rmax达到了0.24 mL·h-1.
2) 0.02%氯仿+腐殖酸系统的液相末端产物以乙醇、乙酸和丁酸为主, 且丙酸含量低于其他发酵系统, 另外, 挥发酸总量显著高于其他组(p < 0.01).
3) 对照组中的优势菌群为Ottowia、Ignavibacterium、Nitrospira、Saccharibacteria_genera_incertae_sedis和Terrimonas;当加入0.02%氯仿和腐殖酸后, Ottowia、Saccharibacteria_genera_incertae_sedis和Ferruginibacter的相对丰度有所增加, 而Nitrospira的相对丰度显著减少.
Abubackar H N, Keskin T, Yazgin O, et al. 2019. Biohydrogen production from autoclaved fruit and vegetable wastes by dry fermentation under thermophilic condition[J]. International Journal of hydrogen Energy, 44: 18776-18784. DOI:10.1016/j.ijhydene.2018.12.068 |
Antwi P, Li J, Boadi P O, et al. 2017. Functional bacterial and archaeal diversity revealed by 16S rRNA gene pyrosequencing during potato starch processing wastewater treatment in an UASB[J]. Bioresource Technology, 235: 348-357. DOI:10.1016/j.biortech.2017.03.141 |
Arimi M M, Knodel J, Kiprop A, et al. 2015. Strategies for improvement of biohydrogen production from organic rich wastewater: A review[J]. Biomass and Bioenergy, 75: 101-118. DOI:10.1016/j.biombioe.2015.02.011 |
班巧英, 岳立峰, 李建政, 等. 2020. 萘厌氧降解菌群的富集及氧化还原介体的强化[J]. 中国环境科学, 40(7): 3150-3155. DOI:10.3969/j.issn.1000-6923.2020.07.042 |
班巧英, 刘琦, 余敏, 等. 2019. 氧化还原介体催化强化污染物厌氧降解研究进展[J]. 科技导报, 37(21): 88-96. |
Fuess L T, Zaiat F M, do Nascimento C A O. 2019. Novel insights on the versatility of biohydrogen production from sugarcane vinasse via thermophilic dark fermentation: Impacts of pH-driven operating strategies on acidogenesis metabolite profiles[J]. Bioresource Technology, 286: 121379. DOI:10.1016/j.biortech.2019.121379 |
国家环保局《水和废水监测分析方法》编委会. 2002. 水和废水监测分析方法[M]. 北京: 中国环境科学出版社, 27-30.
|
金鑫, 符波, 朱慧, 等. 2019. 产甲烷抑制剂氯仿对污泥厌氧消化中同型产乙酸作用的影响[J]. 环境科学学报, 5: 1536-1543. |
Li H B, Lian J, Guo Y K, et al. 2012. Biocatalyst of redox mediators on the denitrification by Paracoccus versutus Strain GW1[J]. Envrironmental Science, 33(7): 2458-2463. |
李海波, 廉静, 郭延凯, 等. 2012. 氧化还原介体催化强化Paracoccus versutus菌株GW1反硝化特性研究[J]. 环境科学, 33(7): 2458-2463. |
Lin Y, Ye G, Ding W, et al. 2020. Niche differentiation of comammox Nitrospira and canonical ammonia oxidizers in soil aggregate fractions following 27-year fertilizations[J]. Agriculture Ecosystems and Environment, 304: 107147. DOI:10.1016/j.agee.2020.107147 |
Liu Z, Niels-Ulrik F, Kajetan V, et al. 2012. Complete genome of Ignavibacterium album, a metabolically versatile, flagellated, facultative anaerobe from the phylum Chlorobi[J]. Frontiers in Microbiology, 3: 185-200. |
Maspolim Y, Zhou Y, Guo C, et al. 2015. Determination of the archaeal and bacterial communities in two-phase and single-stage anaerobic systems by 454 pyrosequencing[J]. Journal of Environmental Science, 36: 121-129. DOI:10.1016/j.jes.2015.02.017 |
Parameswaran P, Torres C I, Kang D W, et al. 2012. The role of homoacetogenic bacteria as efficient hydrogen scavengers in microbial electrochemical cells (MXCs)[J]. Water Science and Technology, 65(1): 1-6. DOI:10.2166/wst.2011.519 |
Qin Y, Li L, Wu J, et al. 2019. Co-production of biohydrogen and biomethane from food waste and paper waste via recirculated two-phase anaerobic digestion process: Bioenergy yields and metabolic distribution[J]. Bioresource Technology, 276: 325-334. DOI:10.1016/j.biortech.2019.01.004 |
Rodrigues C, Núñez-Gómez D, Follmann H V D M, et al. 2020. Biostimulation of sulfate-reducing bacteria and metallic ions removal from coal mine-impacted water (MIW) using shrimp shell as treatment agent[J]. Journal of Hazardous Materials, 398: 122893. DOI:10.1016/j.jhazmat.2020.122893 |
Spring S, Jäckel U, Wagner M, et al. 2004. Ottowia thiooxydans gen. nov., sp. nov., a novel facultatively anaerobic, N2O-producing bacterium isolated from activated sludge, and transfer of Aquaspirillum gracile to Hylemonella gracilis gen. nov., comb. nov[J]. International Journal of Systematicand Evolutionary Microbiology, 54: 99-106. DOI:10.1099/ijs.0.02727-0 |
王晋, 刘和, 许科伟, 等. 2011. 污泥厌氧消化过程中产氢产乙酸/同型产乙酸协同产酸研究[J]. 环境科学, 32(6): 1673-1678. |
Yang G, Wang J. 2019. Biohydrogen production by co-fermentation of sewage sludge and grass residue: Effect of various substrate concentrations[J]. Fuel, 237: 1203-1208. DOI:10.1016/j.fuel.2018.10.026 |
Zhang D, Zhang N, Yu X, et al. 2017. Effect of humins from different sediments on microbial degradation of 2, 2', 4, 4', 5, 5'- hexachlorobiphenyl (PCB153), and their polyphasic characterization[J]. Rsc Advances, 7(12): 6849-6855. |
张立国, 班巧英, 李建政. 2016. UASB反应器中产甲烷菌对温度胁迫的响应[J]. 中国环境科学, 36(4): 1082-1086. |