 2021, Vol. 41
2021, Vol. 41



微囊藻毒素(Microcystins, MCs)是一类由蓝藻产生的单环七肽结构的天然肝毒素, 根据其组成氨基酸的不同而存在多种异构体, 其中, 微囊藻毒素-LR(MC-LR)由于其存在普遍、性质稳定、含量较高、毒性较强受到广泛研究(苏雅玲等, 2013).MCs的人工去除方法主要有物理法、化学法、生物法及其组合方法, 其中, 物理法包括炭吸附、超滤、反渗透等, 化学法包括氯、H2O2、O3等氧化剂氧化和光催化氧化等, 生物法有生物膜反应器、人工湿地系统等(王东伟, 2019).在湖泊水环境治理领域, 化学药剂不适于大量投加到水体中, 会造成二次污染;光催化氧化技术成本较高;生物膜、超滤等方法需要大型设备, 也不适用于湖泊治理.相比之下, 生物炭吸附是相对廉价且实用的方法.
生物炭是生物质在限氧条件下通过热化学转化获得的固体材料(Lehmann et al., 2009; IBI, 2015), 在土壤改良、节能减排及缓解气候变化、水体和土壤污染的修复等方面都具有重要的应用价值, 并在环境保护领域展现出巨大的潜力(Lehmann, 2007; Qian et al., 2015).生物炭因具有较强的吸附能力而被广泛用于水和土壤中有机污染物的去除研究(Zhang et al., 2013).目前制备生物炭的原料有很多, 农林废料、生活垃圾、畜禽粪便、污泥沼渣等均可制备成生物炭(Ahmad et al., 2014).水生植物由于来源广泛、生长迅速、能快速吸收水体中的氮、磷等营养元素, 被广泛应用于污染水体生态修复中, 但其生长具有季节性, 每年须及时收割, 否则会大量腐烂并影响水质造成二次污染(郑孟杰等, 2017).因此, 将水生植物资源化利用是解决这一问题的有效方法.传统的资源化利用途径有制备饲料、肥料、燃料等, 而将水生植物残体作为原料制备生物炭是水生植物资源化利用的新途径, 近年来受到越来越多的关注(杨柳燕等, 2016).
为了提高生物炭的吸附性能, 通常会对生物炭进行改性.生物炭的改性方法主要有蒸汽改性、酸改性、碱改性、氧化剂改性、金属盐/氧化物改性等(Wang et al., 2019).其中, 磷酸改性具有相对较低的热解温度、对设备腐蚀性较小、污染小、成本低等优点(Chu et al., 2018).
目前, 有关生物炭吸附微囊藻毒素的研究较少, 而利用改性水生植物生物炭吸附水体中微囊藻毒素的研究尚未见报道, 有关生物炭对MC-LR的改性和吸附机理也有待进一步深入研究.因此, 本研究选用湖泊、湿地生态系统中常见的两种水生植物苦草(Vallisneria natans)和狐尾藻(Myriophyllum verticillatum L.)为原料, 采用不同热解温度制备生物炭, 并进行磷酸改性, 研究苦草和狐尾藻生物炭及其磷酸改性生物炭对MC-LR的吸附去除性能, 并探讨pH、离子强度和溶解性有机质(DOM)对吸附的影响.
2 材料与方法(Materials and methods) 2.1 材料、试剂和仪器材料:水生植物苦草和狐尾藻采自太湖贡湖湾湿地公园内.
试剂:MC-LR(CAS: 101043-37-2, ≥95% by HPLC)购自Taiwan Algal Science Inc.;微囊藻毒素检测试剂盒购自Beacon Analytical Systems Inc.;其他实验用试剂均为分析纯.
仪器:扫描电子显微镜(FEI QUANTA FEG 250)、有机元素分析仪(Elementar vario EL III)、全自动比表面及孔隙度分析仪(麦克ASAP2460)、X射线光电子能谱仪(Thermo Fisher Nexsa)、傅里叶变换红外光谱仪(NICOLET NEXUS870)、酶标仪(TECAN Infinite 200 PRO)、马弗炉(HY-1000 ℃, 洛阳恒宇实验电炉厂)、pH计(Thermo Orion Star A211)、恒温摇床、超纯水机等.
2.2 生物炭的制备和改性将新鲜植物水洗, 晾干, 再放置于60 ℃鼓风干燥箱烘干至恒重.用粉碎机粉碎过20目筛, 在坩埚中填满, 用马弗炉分别升温至350、500、650 ℃维持3 h, 得到初始生物炭, 苦草生物炭分别标记为K350、K500、K650, 狐尾藻生物炭分别标记为H350、H500、H650.
按照文献中报道的方法对初始生物炭进行磷酸改性(Peng et al., 2017; Chu et al., 2018).将生物炭以浸渍比1:1(生物炭(g): 磷酸溶液(g))浸渍于42.5%的磷酸溶液中24 h, 然后再分别在马弗炉中加热至制备生物炭对应的温度(350、500、650 ℃), 维持2 h.随后多次水洗至中性, 60 ℃烘干, 得到磷酸改性生物炭, 分别标记为K350-P、K500-P、K650-P、H350-P、H500-P、H650-P.
将所有制备得到的生物炭研磨过筛, 取100~200目颗粒并再次水洗, 从而把生物炭的颗粒粒径控制在相同的范围, 烘干后放在干燥器中储存.
2.3 生物炭的表征利用扫描电镜(SEM)观察生物炭的微观结构.用元素分析仪分别测定生物炭中的碳、氢、氮、硫元素含量(CHNS模式)和氧元素含量(O模式).用全自动比表面及孔隙度分析仪测定生物炭在77 K温度下的氮气吸附等温线, 并计算生物炭的比表面积和孔径分布.用X射线光电子能谱(XPS)分析生物炭的表面元素含量和化学形态.利用KCl压片法, 测定生物炭在4000~400 cm-1的傅里叶红外光谱(FTIR).
2.4 吸附实验 2.4.1 吸附去除率测定配制10 μg·L-1的MC-LR溶液, 以0.001 mol·L-1 pH=7.0的磷酸盐缓冲液(0.001 mol·L-1 NaH2PO4溶液并用NaOH调节至pH=7.0)为背景溶液.取20.0 mg生物炭和50 mL上述MC-LR溶液混合, 在25 ℃恒温摇床中振荡24 h, 然后用0.45 μm针头滤膜过滤得到滤液, 测定MC-LR含量.
2.4.2 吸附动力学取4.0 mg生物炭与100 mL上述10 μg·L-1 MC-LR溶液混合, 在25 ℃恒温摇床中振荡, 每间隔一段时间取约2 mL混合液并用0.45 μm针头滤膜过滤, 测定滤液中MC-LR含量.
2.4.3 pH的影响为了探究pH对吸附性能的影响, 配制10 μg·L-1的MC-LR溶液, 以0.001 mol·L-1的磷酸二氢钠溶液为背景缓冲液并分别用H2SO4/NaOH调节pH为4.0、6.0、8.0、10.0.取2.0 mg的生物炭与50 mL上述不同pH的MC-LR溶液混合, 在25 ℃恒温摇床中振荡24 h, 然后用0.45 μm针头滤膜过滤得到滤液, 测定MC-LR含量.
2.4.4 离子强度的影响由于Na2SO4在水溶液中完全电离、不会发生水解的特性, 选用Na2SO4作为无机电解质调节离子强度.配制10 μg·L-1的MC-LR溶液, 以0.001 mol·L-1 pH=7.0的磷酸缓冲液为背景溶液, 添加Na2SO4, 使溶液中Na2SO4的浓度分别为0、0.02、0.05、0.10 mol·L-1.取2.0 mg生物炭与50 mL上述不同离子强度的MC-LR溶液混合, 在25 ℃恒温摇床中振荡24 h, 然后用0.45 μm针头滤膜过滤得到滤液, 测定MC-LR含量.
2.4.5 DOM的影响为了测定DOM对生物炭吸附MC-LR性能的影响, 分别选用单宁酸(CAS: 1401-55-4)、富里酸(CAS: 479-66-3)、没食子酸(CAS: 149-91-7)作为模式DOM.配制10 μg·L-1的MC-LR溶液, 以0.001 mol·L-1 pH=7.0的磷酸缓冲液为背景溶液, 分别添加DOM, 使溶液中DOM的浓度为2.0 μmol·L-1.取2.0 mg生物炭与50 mL上述含有不同DOM的MC-LR溶液混合, 在25 ℃恒温摇床中振荡24 h, 然后用0.45 μm针头滤膜过滤得到滤液, 测定MC-LR含量.
2.5 MC-LR的测定利用酶联免疫吸附法测定溶液中的微囊藻毒素浓度.实验用微囊藻毒素检测试剂盒的量程为0.1~2.0 μg·L-1, 按照说明书上的步骤进行操作, 用酶标仪读板, 然后绘制标准曲线即可计算出样品中MC-LR的浓度.超过量程的样品需要经过适当比例稀释.对于酸性或碱性较强的样品, 利用pH=7.0的磷酸盐缓冲液稀释后才能进行测定.吸附实验中所有实验都进行平行样测定.
3 结果与讨论(Results and discussion) 3.1 不同温度制备生物炭的得炭率得炭率是植物热解后炭的质量与热解前植物干重的比值.从图 1可以看出, 随着热解温度的升高, 得炭率越来越低.这是因为随着温度的升高, 植物中的有机物逐渐分解、气化, 导致质量减少.研究表明, 当热解温度处于较低的350 ℃时, 主要发生结合水的脱除和纤维素、半纤维素分解;热解温度升高到500 ℃以上时, 纤维素已基本完全分解, 稳定性较强的木质素则缓慢分解, 热解过程中有机物会分解成H2、CO、CO2及一些碳氢化合物;当温度进一步升高接近700 ℃时, 基本完全炭化(Yang et al., 2007; Xiao et al., 2014).另外, 结果显示由狐尾藻制备生物炭的得炭率要略高于苦草.
 |
| 图 1 苦草和狐尾藻在不同温度下制备生物炭的得炭率 Fig. 1 Carbon yield rate of Vallisneria and Myriophyllum biochar at different temperatures |
生物炭及磷酸改性生物炭的扫描电镜图见图 2, 放大倍数为1100倍.从图中可以看出, 较高温度(500、650 ℃)制备的苦草和狐尾藻生物炭表面孔更加密集, 粗糙程度也要明显高于较低温度下(350 ℃)制备的生物炭.磷酸改性也能够增加生物炭表面的粗糙程度, 尤其是较低温度(350 ℃)下制备的生物炭.
 |
| 图 2 生物炭和磷酸改性生物炭的SEM图 Fig. 2 SEM of biochar and phosphoric acid modified biochar |
基于元素分析仪的结果和XPS的结果, 表 1中列了生物炭及磷酸改性生物炭中元素碳、氢、氧、氮的比例和表面元素磷的比例, 以及O/C、(N+O)/C、H/C的计算结果.O/C、(N+O)/C比值越大, 说明生物炭的极性越强;H/C比值越小, 说明芳香性越强(富丽等, 2019).结果表明, 未经改性的生物炭中, 较低温度下(350 ℃)制备的生物炭具有更高的含氧量且极性更强;而随着温度的升高, 含氧量降低, 但芳香性增强.此外, 磷酸改性增加了生物炭表面磷元素的含量, 且越高温度下制备的磷酸改性生物炭的磷含量越高.
| 表 1 生物炭的元素组成和比例 Table 1 Elemental composition and ratio of biochar |
基于生物炭在77 K下N2吸附等温线, 用BET法计算生物炭的比表面积SBET, 用t-plot法计算生物炭的微孔比表面积Smic, 总孔容Vtot由等温线p/p0=0.99处N2的吸附量计算得出, 平均孔径Davg是由4Vtot/SBET计算得到.利用DFT模型得出的孔径分布算出微孔孔容Vmic(孔径≤2 nm)和中孔孔容Vmeso(2 nm<孔径≤50 nm).本研究生物炭的比表面和孔径分析结果见表 2.
| 表 2 生物炭的比表面和孔径分析 Table 2 Specific surface and pore size analysis of biochar |
从表 2中可以看出, 随着制备温度的提高, SBET明显升高, 尤其在650 ℃下制备的生物炭比表面积远远大于350 ℃和500 ℃下制备的生物炭比表面积.高温制备的生物炭有更小的平均孔径和更大的孔体积, 而且微孔体积的增大尤为明显, 如H650的微孔体积是H350的371倍.研究表明, 更高的热解温度可以促进生物炭孔隙的生成, 从而产生更大的比表面积和孔容(Ahmad et al., 2014).磷酸改性对生物炭比表面的影响也十分明显, 可增大350 ℃和500 ℃下制备的生物炭的比表面积.前人的研究指出, 磷酸改性可以促进生物炭多孔结构的生成, 磷酸解离出的氢离子可以促进反应中键的断裂, 还可以通过环化、缩合并形成磷酸盐和多聚磷酸盐, 促进交联, 通过洗涤去除残余的多聚磷酸盐, 就会留下孔隙(Sych et al., 2012).此外, 磷酸和多聚磷酸能够促进生物质中有机质脱水炭化, 脱水产生的蒸汽也能促进孔隙的生成(Jia et al., 2017; Chu et al., 2018).但对于650 ℃下制备的狐尾藻炭(H650), 磷酸改性使比表面积略有降低, 从322 m2·g-1降到281 m2·g-1.已有研究表明, 在磷酸存在时, 温度对孔隙形成的影响减弱, 甚至较高温度下(650 ℃)制备的磷酸改性生物炭的比表面积还会略低于较低温度下制备的改性生物炭(Zhao et al., 2017).
3.2.4 表面基团和化学键FTIR谱图见图 3.波数3200~3400 cm-1处的宽峰是O—H基团伸缩振动(Puziy et al., 2002).2925 cm-1处是脂肪族CH2伸缩振动(富丽等, 2019), 且高温制备的生物炭没有此峰, 说明随着温度的升高, 脂肪族CH2减少.1694、1727 cm-1处的小峰是羰基C=O伸缩振动(Ahmed et al., 2018).1578~1615 cm-1处是芳香族C=C、C=O伸缩振动(杨雪等, 2018).1432~1445 cm-1处的小峰是脂肪族CH2的变形振动(Zheng et al., 2013; 陈翠苹, 2016).1420~1431 cm-1处的强峰是碳酸根反对称伸缩振动(翁诗甫, 2010), 且低温(350 ℃)制备的生物炭和磷酸改性的生物炭中没有出现此峰.1315~1316 cm-1处是CH2扭曲振动(翁诗甫, 2010).1260~1288 cm-1处是酚类C—OH伸缩振动(翁诗甫, 2010).1016~1058 cm-1处是C—O—C伸缩振动(Ahmed et al., 2018).875、746、712 cm-1处的多个吸收峰是芳环C—H面外变形振动(Puziy et al., 2002; Yang et al., 2014).772~781 cm-1处则是二氧化硅引起的(Xiao et al., 2014).磷酸改性使生物炭增加了一些含磷基团, 1094~1182 cm-1处是P—O—C和P=OOH中P=O和P—O的伸缩振动(Kennedy et al., 2004; 钟晓晓等, 2018).900~926 cm-1处是焦磷酸盐中P—O—P不对称伸缩振动(Puziy et al., 2002).
 |
| 图 3 生物炭的FTIR图 Fig. 3 FTIR of biochar |
对生物炭进行XPS分析, 以苦草炭K350、K650和K350-P为例(图 4).图 4a是K350的XPS全扫图, 结果表明, 生物炭中除C、N、O元素外, 还存在少量的Si、Al、Ca等矿物元素.磷酸改性后, 出现P2p的峰(图 4b).对C1s谱峰用XPS分析软件(Avantage 5.967)进行分峰, 图 4c是K350的C1s谱峰及其分峰拟合, 284.5 eV附近的是C—C峰, 286.0 eV处是C—O(醇), 286.9~287.0 eV处是C—O—C(醚), 287.8~288.0 eV处是C=O(酮、醛), 288.6~288.8 eV处是O—C=O(羧基)(Moulder et al., 1995; Peng et al., 2017).高温制备的生物炭(K650)还会形成π-π*峰(图 4d), 这是芳环中离域电子导致的(Fan et al., 2010; Li et al., 2018a).磷酸改性生物炭(K350-P)的C1s谱峰及分峰结果与K350相似(图 4e).图 4f显示, 磷酸改性生物炭(K350-P)有明显的P2p谱峰, 而未改性生物炭无明显P2p谱峰.P2p3/2轨道峰在133.3 eV处, 说明磷的化学形态主要是焦磷酸盐(Moulder et al., 1995).FTIR和XPS分析结果表明, 生物炭中存在丰富的含氧基团.生物炭表面丰富的含氧官能团能与亲水性MC-LR中的羧基及氨基等形成氢键, 有利于MC-LR在生物炭表面上的吸附.
 |
| 图 4 生物炭的XPS谱图 (a.K350的XPS全扫图, b. K350-P的XPS全扫图, c. K350的C1s谱及其分峰拟合图, d.K650的C1s谱及其分峰拟合图, e. K350-P的C1s谱及其分峰拟合图, f. K350的P2p谱和K350-P的P2p谱及其分峰拟合图) Fig. 4 XPS of biochar (a. XPS survey spectrum of K350, b. XPS survey spectrum of K350-P, c. XPS spectrum and fitting curves of C1s for K350, d. XPS spectrum and fitting curves of C1s for K650, e. XPS spectrum and fitting curves of C1s for K350-P, f. The P2p spectra of K350 and K350-P) |
不同温度制备的苦草和狐尾藻生物炭对溶液中MC-LR的去除率结果见图 5a, 图中K表示苦草生物炭, H表示狐尾藻生物炭.可以看出, 随着热解温度的提高, 去除率也明显增加, 350 ℃制备的生物炭对MC-LR去除率不到20%, 而650 ℃制备的生物炭对MC-LR的去除率接近100%.可见生物炭对MC-LR的吸附能力也随着制备热解温度的提高而提高.根据生物炭比表面的表征结果, 更高温度制备的生物炭具有更大的比表面积和孔容, 从而有利于MC-LR的吸附.此外, 在较低热解温度下, 不完全碳化可能使大多数无定形碳保留在生物炭上, 而更高的热解温度会去除残留的脂肪族有机物以形成更多的孔, 并将无定形碳转化为较稠密的芳香族碳(Keiluweit et al., 2010).高芳香性碳表面可以作为π电子供体, 而MC-LR的胍基存在共轭效应, 可以作为π电子受体, 于是高温制备的生物炭与MC-LR之间会产生π+-π电子供体-受体相互作用增强吸附(Li et al., 2018b).这也可以解释650 ℃热解制备的苦草和狐尾藻生物炭具有较高的吸附能力.
 |
| 图 5 苦草(K)和狐尾藻(H)生物炭对MC-LR的去除率 (a.未改性生物炭(炭投加量20 mg/50 mL), b.磷酸改性生物炭(炭投加量20 mg/50 mL), c.磷酸改性生物炭(炭投加量2.0 mg/50 mL)) Fig. 5 Removal rate of MC-LR by Vallisneria (K) and Myriophyllum (H) biochar (a. original biochar (biochar addition: 20 mg/50 mL), b. phosphoric acid modified biochar (biochar addition: 20 mg/50 mL), c. phosphoric acid modified biochar (biochar addition: 2.0 mg/50 mL)) |
图 5b显示了磷酸改性苦草和狐尾藻生物炭对溶液中MC-LR的去除率结果.可以看出, 与未经改性的生物炭相比, 磷酸改性使生物炭对藻毒素的吸附能力明显提高, 尤其是350 ℃制备的生物炭, 磷酸改性使其对MC-LR去除率提高到95%以上.根据生物炭比表面的表征结果, 磷酸改性促进了孔隙的生成, 孔隙的增加有利于吸附MC-LR.研究表明, 孔填充作用是生物炭吸附MC-LR的重要机制, MC-LR分子容易被吸附到尺寸相似的孔隙中(Li et al., 2018b; 潘禹等, 2018).MC-LR分子尺寸为1.9 nm×1.5 nm×1.1 nm(Lanaras et al., 1991; Pendleton et al., 2001), 结合表 2数据, 高温(650 ℃)制备生物炭及磷酸改性的生物炭其平均孔径(2~4 nm)略大于MC-LR分子, 有利于对MC-LR的吸附.
由于图 5b中各磷酸改性生物炭对MC-LR去除率接近100%, 故难以相互区分出差别.为了比较各磷酸改性生物炭对MC-LR的吸附能力, 减少吸附体系中炭的投加量, 使用2.0 mg/50 mL炭投加量进行去除率实验, 结果见图 5c.可以发现, 不同温度制备的磷酸改性生物炭, 去除率并没有明显差异.
3.4 吸附动力学本文进行了K650、K650-P、H650和H650-P的吸附动力学实验, 采用公式(1)计算吸附量qt, 再利用OriginPro 8.5软件进行吸附准一级动力学方程(式(2))、准二级动力学方程(式(3))和颗粒内扩散方程(式(4))拟合(Ho et al., 1999a; Kumar, 2007; 郎印海等, 2014).
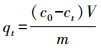
|
(1) |
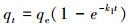
|
(2) |
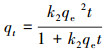
|
(3) |
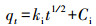
|
(4) |
式中, t为吸附时间(h), qt为t时刻吸附量(μg·g-1), c0为MC-LR初始浓度(μg·L-1), ct为t时刻MC-LR浓度(μg·L-1), V为溶液体积(L), m为炭投加量(g), 以上指标均为实验设定或者测量所得.其余指标qe、k1、k2、ki、Ci是公式中拟合计算出的参数, 其中, qe为计算出的平衡吸附量(μg·g-1).
拟合计算参数见表 3, 拟合结果见图 6.从图 6中可以看出, 各生物炭对MC-LR的吸附速率都呈现出初始阶段吸附速率较快, 随着时间的推移, 吸附速率减慢的特征.这可能是因为随着吸附的进行, 生物炭表面的吸附位点和孔隙被吸附质逐渐占据, 吸附位点减少且排斥阻力增加(Li et al., 2018a);此外, 溶液中MC-LR的浓度也随着吸附的进行而降低, 使扩散驱动力减弱, 也会导致吸附速率减慢(刘倩辉等, 2010).
| 表 3 生物炭和磷酸改性生物炭吸附动力学参数 Table 3 Adsorption kinetic parameters of biochar and phosphoric acid modified biochar |
 |
| 图 6 生物炭和磷酸改性生物炭吸附动力学方程拟合 Fig. 6 Fitting kinetic equations of biochar and phosphoric acid modified biochar adsorption |
图 6显示3种动力学方程的拟合曲线均能表现出较好的拟合效果.从表 3中可以看出, 准一级动力学方程对狐尾藻生物炭(H650、H650-P)的拟合优度(R2)比苦草生物炭(K650、K650-P)要好.准一级动力学方程表示反应速率与反应物浓度呈正比, 一般适合描述边界液膜扩散机制(Ho et al., 1999b).对于K650、K650-P、H650, 准二级动力学方程的拟合优度要高于准一级动力学方程, 这表明吸附过程的速率可能受到化学吸附作用的控制, 即吸附质和吸附剂之间存在共享或交换电子的化学作用(Hameed et al., 2008).准一级动力学和准二级动力学的qe值表明磷酸活化生物炭对MC-LR的平衡吸附量要高于未经活化生物炭的吸附量.颗粒内扩散模型对K650-P、H650也表现出较好的拟合优度, 且Ci值不为0表明颗粒内扩散并非唯一的速率控制步骤, 边界扩散在一定程度上也影响了吸附(Cheung et al., 2007).综上分析, 生物炭和磷酸改性生物炭对MC-LR的吸附是多种因素共同作用的结果.
3.5 pH的影响在不同pH条件下进行K650、K650-P、H650和H650-P对MC-LR的去除实验, 结果见图 7.从图中可以看出, 随着pH的增大, 生物炭对MC-LR的去除率呈下降趋势, 这可能和MC-LR与生物炭之间的静电作用力有关.研究表明, MC-LR表面带有的羧基和氨基会在不同pH条件下发生电离(pKa1=2.09, pKa2=2.19, pKa3=12.48), 从而形成MCLR+、MCLR0、MCLR-、MCLR2- 4种形态(Maagd et al., 1999; Li et al., 2014).根据其pKa值可计算出各形态在不同pH条件下的百分比(图 8), 当溶液pH为4~10时, MC-LR是以MCLR-的形态存在.溶液pH也会影响生物炭的带电情况, 当溶液pH高于生物炭的零电荷点pH(pHpzc)时, 生物炭表面带负电, 且pH越高所带负电荷越多, 从而对MCLR-的静电排斥作用越强;反之若溶液pH低于生物炭的pHpzc时, 生物炭表面带正电, 则pH越低产生的静电吸引作用越强(Li et al., 2018b).因此, 溶液pH越大, 生物炭对MC-LR的吸附性能越差.从图 7中还可以看出, pH对苦草生物炭吸附性能的影响较狐尾藻生物炭小.此外, 磷酸改性生物炭对MC-LR的去除率随着pH增大的下降趋势要比未改性生物炭的下降趋势小, 说明磷酸改性能减弱pH对生物炭吸附MC-LR效率的影响.同时进行了pH=4~10下的空白对照实验(数据未显示), 结果表明, MC-LR在不加生物炭水中24 h的浓度基本没有变化.
 |
| 图 7 生物炭和磷酸改性生物炭在不同pH条件下对MC-LR的去除率 Fig. 7 Removal rate of biochar and phosphoric acid modified biochar at different pH |
 |
| 图 8 各pH条件下MC-LR形态分布图 Fig. 8 Distribution of MC-LR species at various pH conditions |
在不同离子强度下进行K650、K650-P、H650和H650-P对MC-LR的去除实验, 采用Na2SO4作调节离子强度的电解质, 结果见图 9.从图中可以看出, K650受离子强度的影响较大, 表现为电解质的存在抑制了吸附.这可能是因为溶液中的离子减弱了静电相互作用(Campinas et al., 2006), 从而减小了生物炭与MC-LR之间的静电引力使吸附性能减弱.也有可能是因为电解质与吸附质离子之间发生离子交换, 产生的竞争作用使生物炭对MC-LR的吸附量减少(吴志坚等, 2010), 这也间接表明阴离子交换作用可能是导致生物炭吸附MC-LR的因素之一(Li et al., 2014).不过, 实验结果中随着离子强度的增大, K650对MC-LR的去除率没有出现进一步下降的趋势, 表明离子强度对吸附的影响十分有限.而且离子强度似乎对K650-P、H650、H650-P的吸附性能并没有影响, 这也说明对磷酸改性生物炭和狐尾藻生物炭而言, 阴离子交换的作用微不足道.
 |
| 图 9 生物炭和磷酸改性生物炭在不同离子强度下对MC-LR的去除率 Fig. 9 Removal rate of biochar and phosphoric acid modified biochar at different ionic strength |
考虑到天然水体中含有各类溶解性有机质(DOM), 而这些DOM可能会影响生物炭吸附MC-LR, 本实验选用单宁酸(TA)、富里酸(FA)、没食子酸(GA)作为模式DOM, 这3种有机物的分子量(MW)分别为1701.2、308.24、170.12 g·mol-1, 实验结果见图 10.从图中可以看出, GA对生物炭吸附性能的影响较小, 而TA则能明显降低MC-LR的吸附去除率, 其规律表现为分子量越大, 对生物炭的吸附抑制效果也越明显.DOM可以通过竞争和堵塞孔从而降低生物炭对MC-LR的吸附效率(刘倩辉等, 2010).研究表明, MC-LR由于其分子量(MW: 995.2)和分子尺寸(1.9 nm×1.5 nm×1.1 nm)较大, 主要通过生物炭中孔(孔径2~50 nm)进行吸附(Li et al., 2018b), 因此, 同样是较大分子量的TA与之竞争作用更为明显;而分子量较小的GA主要占据微孔(Xiao et al., 2012), 不会产生强烈的竞争作用.此外, TA(1.93 nm×1.73 nm×1.32 nm)比MC-LR有着更大的体积(Xiao et al., 2012), 除了竞争作用, 还能够产生堵塞作用阻止MC-LR进入合适大小的孔隙.FA分子量介于TA和GA之间, 则对生物炭吸附性能的影响也居中.这些结果间接表明生物炭吸附MC-LR存在孔填充作用机制.实验还表明, 磷酸改性能使DOM对生物炭吸附的抑制作用减小, 因而有利于在DOM存在的条件下吸附水中MC-LR.
 |
| 图 10 生物炭和磷酸改性生物炭在不同DOM下对MC-LR的去除率 Fig. 10 Removal rate of biochar and phosphoric acid modified biochar at different DOMs |
1) 高温(650 ℃)热解制备的苦草和狐尾藻生物炭对MC-LR有较好的去除性能, 磷酸改性能够进一步提高生物炭的吸附性能.因此, 水生植物生物炭可以用于吸附水中MC-LR, 为减少水体微囊藻毒素污染危害提供新的解决方案, 有良好的环境效益和应用前景.
2) 热解温度越高, 生物炭得炭率越低.高温制备的生物炭及磷酸改性的生物炭有着较大的比表面积和较多的微孔, 平均孔径在2~4 nm.生物炭表面有丰富的含氧官能团, 磷酸改性生物炭表面增加了含磷官能团.
3) 生物炭对MC-LR的吸附速率随着时间的推移而减慢.准一级动力学方程、准二级动力学方程和颗粒内扩散方程均能表现出较好的拟合效果, 说明生物炭和磷酸改性生物炭对MC-LR的吸附存在多种因素共同作用, 包括液膜扩散、化学作用、颗粒内扩散机理.
4) 溶液pH和DOM的存在会影响生物炭对MC-LR的吸附性能, 表现为pH越高、DOM分子量越大, MC-LR的吸附去除率越低, 但离子强度对生物炭的吸附性能影响不大;而磷酸改性能减弱pH和DOM对生物炭吸附MC-LR效率的影响.推测生物炭吸附MC-LR可能存在氢键、静电作用和孔填充作用.
Ahmad M, Rajapaksha A U, Lim J E, et al. 2014. Biochar as a sorbent for contaminant management in soil and water: A review[J]. Chemosphere, 99: 19-33. DOI:10.1016/j.chemosphere.2013.10.071 |
Ahmed M J, Hameed B H. 2018. Adsorption behavior of salicylic acid on biochar as derived from the thermal pyrolysis of barley straws[J]. Journal of Cleaner Production, 195: 1162-1169. DOI:10.1016/j.jclepro.2018.05.257 |
Campinas M, Rosa M J. 2006. The ionic strength effect on microcystin and natural organic matter surrogate adsorption onto PAC[J]. Journal of Colloid and Interface Science, 299(2): 520-529. DOI:10.1016/j.jcis.2006.02.042 |
陈翠苹. 2016. 水中亚硝胺类污染物的吸附去除与作用机制[D]. 杭州: 浙江大学. 16-33
|
Cheung W H, Szeto Y S, McKay G. 2007. Intraparticle diffusion processes during acid dye adsorption onto chitosan[J]. Bioresource Technology, 98(15): 2897-2904. DOI:10.1016/j.biortech.2006.09.045 |
Chu G, Zhao J, Huang Y, et al. 2018. Phosphoric acid pretreatment enhances the specific surface areas of biochars by generation of micropores[J]. Environmental Pollution, 240: 1-9. DOI:10.1016/j.envpol.2018.04.003 |
Fan Y, Wang B, Yuan S H, et al. 2010. Adsorptive removal of chloramphenicol from wastewater by NaOH modified bamboo charcoal[J]. Bioresource Technology, 101(19): 7661-7664. DOI:10.1016/j.biortech.2010.04.046 |
富丽, 徐先英, 付贵全, 等. 2019. 五种生物质炭的特性分析[J]. 干旱区资源与环境, 33(9): 202-208. |
Hameed B H, Tan I A W, Ahmad A L. 2008. Adsorption isotherm, kinetic modeling and mechanism of 2, 4, 6-trichlorophenol on coconut husk-based activated carbon[J]. Chemical Engineering Journal, 144(2): 235-244. DOI:10.1016/j.cej.2008.01.028 |
Ho Y S, McKay G. 1999a. A kinetic study of dye sorption by biosorbent waste product pith[J]. Resources, Conservation & Recycling, 25(3): 171-193. |
Ho Y S, McKay G. 1999b. Pseudo-second order model for sorption processes[J]. Process Biochemistry, 34(5): 451-465. DOI:10.1016/S0032-9592(98)00112-5 |
IBI. 2015. Standardized product definition and product testing guidelines for biochar that is used in soil[R]. Canandaigua: International Biochar Initiative
|
Jia Y L, Lu Y, Zhang G X, et al. 2017. Facile synthesis of an eco-friendly nitrogen-phosphorus ammonium salt to enhance the durability and flame retardancy of cotton[J]. Journal of Materials Chemistry A, 5(20): 9970-9981. DOI:10.1039/C7TA01106G |
Keiluweit M, Nico P S, Johnson M G, et al. 2010. Dynamic molecular structure of plant biomass-derived black carbon (biochar)[J]. Environmental Science & Technology, 44(4): 1247-1253. |
Kennedy L J, Vijaya J J, Sekaran G. 2004. Effect of two-stage process on the preparation and characterization of porous carbon composite from rice husk by phosphoric acid activation[J]. Industrial & Engineering Chemistry Research, 43(8): 1832-1838. |
Kumar K V. 2007. Pseudo-second order models for the adsorption of safranin onto activated carbon: Comparison of linear and non-linear regression methods[J]. Journal of Hazardous Materials, 142(1/2): 564-567. |
Lanaras T, Cook C M, Eriksson J E, et al. 1991. Computer modelling of the 3-dimensional structures of the cyanobacterial hepatotoxins microcystin-LR and nodularin[J]. Toxicon, 29(7): 901-906. DOI:10.1016/0041-0101(91)90228-J |
郎印海, 刘伟, 王慧. 2014. 生物炭对水中五氯酚的吸附性能研究[J]. 中国环境科学, 34(8): 2017-2023. |
Lehmann J. 2007. A handful of carbon[J]. Nature, 447: 143-144. DOI:10.1038/447143a |
Lehmann J, Joseph S. 2009. Biochar for Environmental Management Science and Technology[M]. London, UK: Earthscan, 1-12.
|
Li Y R, Zhang J, Liu H. 2018a. Removal of chloramphenicol from aqueous solution using low-cost activated carbon prepared from typha orientalis[J]. Water, 10(4): 351. DOI:10.3390/w10040351 |
Li J M, Cao L R, Yuan Y, et al. 2018b. Comparative study for microcystin-LR sorption onto biochars produced from various plant- and animal-wastes at different pyrolysis temperatures: Influencing mechanisms of biochar properties[J]. Bioresource Technology, 247: 794-803. DOI:10.1016/j.biortech.2017.09.120 |
Li L, Qiu Y P, Huang J X, et al. 2014. Mechanisms and factors influencing adsorption of Microcystin-LR on biochars[J]. Water, Air, & Soil Pollution, 225(12): 2220. DOI:10.1007/s11270-014-2220-6 |
刘倩辉, 裴海燕, 胡文容, 等. 2010. 活性炭去除水体中微囊藻毒素的研究进展[J]. 工业水处理, 30(11): 8-11. |
Maagd P G D, Hendriks A J, Seinen W, et al. 1999. pH-Dependent hydrophobicity of the cyanobacteria toxin microcystin-LR[J]. Water Research, 33(3): 677-680. DOI:10.1016/S0043-1354(98)00258-9 |
Moulder J F, Stickle W F, Sobol P E, et al. 1995. Handbook of X-ray Photoelecctron Spectroscopy[M]. Minnesota: Physical Electronics, Inc.40-41, 40-41, 58-59.
|
潘禹, 王华生, 刘祖文, 等. 2018. 微囊藻毒素活性炭吸附技术研究进展[J]. 水处理技术, 44(8): 6-11, 16. |
Pendleton P, Schumann R, Wong S H. 2001. Microcystin-LR adsorption by activated carbon[J]. Journal of Colloid and Interface Science, 240(1): 1-8. DOI:10.1006/jcis.2001.7616 |
Peng H B, Gao P, Chu G, et al. 2017. Enhanced adsorption of Cu(Ⅱ) and Cd(Ⅱ) by phosphoric acid-modified biochars[J]. Environmental Pollution, 229: 846-853. DOI:10.1016/j.envpol.2017.07.004 |
Puziy A M, Poddubnaya O I, Martínez-Alonso A, et al. 2002. Synthetic carbons activated with phosphoric acid I.Surface chemistry and ion binding properties[J]. Carbon, 40(9): 1493-1505. DOI:10.1016/S0008-6223(01)00317-7 |
Qian K Z, Kumar A, Zhang H L, et al. 2015. Recent advances in utilization of biochar[J]. Renewable and Sustainable Energy Reviews, 42: 1055-1064. DOI:10.1016/j.rser.2014.10.074 |
苏雅玲, 邓一荣. 2013. 富营养化湖泊中微囊藻毒素及其控制去除技术[J]. 环境科学与技术, 36(6): 62-66. DOI:10.3969/j.issn.1003-6504.2013.06.013 |
Sych N V, Trofymenko S I, Poddubnaya O I, et al. 2012. Porous structure and surface chemistry of phosphoric acid activated carbon from corncob[J]. Applied Surface Science, 261: 75-82. DOI:10.1016/j.apsusc.2012.07.084 |
王东伟. 2019. 水环境中微囊藻毒素去除方法的研究进展[J]. 黑龙江环境通报, 43(1): 40-43. |
Wang J L, Wang S Z. 2019. Preparation, modification and environmental application of biochar: A review[J]. Journal of Cleaner Production, 227: 1002-1022. DOI:10.1016/j.jclepro.2019.04.282 |
翁诗甫. 2010. 傅里叶变换红外光谱分析[M]. 北京: 化学工业出版社, 377-388.
|
吴志坚, 刘海宁, 张慧芳. 2010. 离子强度对吸附影响机理的研究进展[J]. 环境化学, 29(6): 997-1003. |
Xiao X Y, Li F L, Huang J X, et al. 2012. Reduced adsorption of propanil to black carbon: Effect of dissolved organic matter loading mode and molecule size[J]. Environmental Toxicology and Chemistry, 31(6): 1187-1193. DOI:10.1002/etc.1800 |
Xiao X, Chen B L, Zhu L Z. 2014. Transformation, morphology, and dissolution of silicon and carbon in rice straw-derived biochars under different pyrolytic temperatures[J]. Environmental Science & Technology, 48(6): 3411-3419. |
Yang G X, Jiang H. 2014. Amino modification of biochar for enhanced adsorption of copper ions from synthetic wastewater[J]. Water Research, 48: 396-405. DOI:10.1016/j.watres.2013.09.050 |
Yang H P, Yan R, Chen H P, et al. 2007. Characteristics of hemicellulose, cellulose and lignin pyrolysis[J]. Fuel, 86(12/13): 1781-1788. |
杨柳燕, 张文, 陈乾坤, 等. 2016. 大型水生植物的资源化利用[J]. 水资源保护, 32(5): 5-10. |
杨雪, 张士秋, 侯其东, 等. 2018. 生物炭的制备及其镁改性对污染物的吸附行为研究[J]. 环境科学学报, 38(10): 4032-4043. |
Zhang X K, Wang H L, He L Z, et al. 2013. Using biochar for remediation of soils contaminated with heavy metals and organic pollutants[J]. Environmental Science and Pollution Research, 20(12): 8472-8483. DOI:10.1007/s11356-013-1659-0 |
Zhao L, Zheng W, Mašek O, et al. 2017. Roles of phosphoric acid in biochar formation: Synchronously improving carbon retention and sorption capacity[J]. Journal of Environmental Quality, 46(2): 393-401. DOI:10.2134/jeq2016.09.0344 |
Zheng H, Wang Z Y, Zhao J, et al. 2013. Sorption of antibiotic sulfamethoxazole varies with biochars produced at different temperatures[J]. Environmental Pollution, 181: 60-67. DOI:10.1016/j.envpol.2013.05.056 |
郑孟杰, 李继洲, 靳红梅, 等. 2017. 沉水植物生物炭对Cr6+和磷的吸附特性[J]. 生态与农村环境学报, 33(12): 1132-1139. DOI:10.11934/j.issn.1673-4831.2017.12.010 |
钟晓晓, 王涛, 王凯, 等. 2018. 磷酸和热解温度对生物炭结构和性质的影响[J]. 湖南师范大学自然科学学报, 41(1): 48-53. |