 2021, Vol. 41
2021, Vol. 41



近年来, 我国海水循环水养殖业发展迅速, 逐渐成为海产品的重要养殖模式(高怡菲等, 2020).海水养殖循环水水量大, 生物代谢产物与剩余饵料分解导致氮、磷等污染物超标, 但营养负荷低, 碳氮比较低且波动大(1~10), 尤其是海水盐效应造成亚硝酸盐累积, 影响了循环利用并增加了水处理排放的难度(Luo et al., 2013;Li et al., 2013;王一笑, 2017).大量未经彻底处理的海水养殖循环水排放可引发近岸海域水体富营养化等系列生态风险(Cao et al., 2007).目前, 国内外学者已对海水养殖废水处理进行了大量研究, 例如, 通过臭氧氧化法以灭活鱼类病原体, 氧化亚硝酸盐和有机废物(包括颜色)(Summerfelt, 2003), 以及通过增强硝化/反硝化过程来强化废水中氮的去除(郑冰冰等, 2020).上述工艺对海水养殖废水具有一定的处理效果, 但仍存在产生二次污染物及废弃污泥难以处理等问题有待解决.
适盐微藻通过光合作用可将无机营养盐、CO2等物质转化成蛋白质、多糖和油脂等生物质, 能够适应低营养海水养殖循环水且可产生生物质资源(Anto et al., 2020).Ansari等(2020)研究发现, 栅藻、小球藻和纤维藻能够去除水产养殖废水中42%~69%的COD和86.45%~98.21%的氨氮.微藻膜反应器处理海水养殖废水能够实现一定的氮、磷去除, 然而出水满足现有《海水养殖水排放要求》(SC/T 9103—2007)仍存在困难, 且亚硝酸盐累积浓度接近1 mg·L-1(马航等, 2019).同时, 微藻水处理应用还面临微藻采收困难的问题.研究发现, 膜光合生物反应器(MPBR)能够实现微藻生物质截留, 并可保证出水浊度达标(陈凡雨等, 2020).对于传统的膜组件聚偏二氟乙烯(PVDF)膜, 由于膜污染导致膜通量下降和能耗增加, 因此, 在微藻采收中不同膜材料性能及膜污染研究十分活跃.活性污泥等混合微生物菌群可利用无纺布形成动态生物膜, 兼具污泥截留和一定的生物降解作用, 比PVDF膜成本更低且易于清洗, 但动态生物膜成膜时间较长(姚嘉等, 2010).同时, 海水养殖循环水的低营养和盐胁迫会抑制微生物生长, 而微藻培养体系微生物群系结构简单, 因此, 有关动态生物膜的形成及效能仍有待探讨.
前期研究发现, 钝顶螺旋藻属于混合营养型, 具有氮同化能力和盐碱耐受性, 碳源充足时主要进行异养代谢合成, 碳源不足时进行自养代谢合成, 能将碳、氮、磷等营养元素转化为生物质资源, 且未发现亚硝酸盐累积(Zhou et al., 2017;刘婷等, 2018).这表明其可能在低碳氮比的海水养殖循环水处理及资源回收中具有应用潜力.
基于此, 本文通过批次实验和膜光合反应器试验研究低碳氮比下碳氮比变化等对钝顶螺旋藻处理海水养殖循环水的性能及生物质产量的影响, 考察PVDF膜和无纺布膜组件构建的膜光合反应器性能及生物质产量, 以期为海水养殖循环水处理及资源化利用提供理论依据.
2 材料与方法(Materials and methods) 2.1 实验废水水质本研究处理的目标废水是海水养殖循环水, 模拟山东沿海某海水鲑鱼养殖基地废水成分如下: 海水素30 g·L-1的人工海水, COD、TP、NH4+-N、NO3--N和NO2--N(分别由葡萄糖、KH2PO4、NH4Cl、NaNO3、NaNO2配制)浓度分别为50、1.16、6.25、3.04和0.51 mg·L-1.在批次实验研究有机碳浓度对微藻处理性能的影响时, 固定氮和磷的浓度, 仅增加有机碳浓度;研究氮浓度影响时, 固定有机碳浓度和氮磷比, 改变氮浓度和磷浓度.
2.2 批次实验设计在装有250 mL模拟海水养殖循环水的500 mL锥形瓶中进行批次试验.接种时将处于指数生长期的钝顶螺旋藻(刘婷等, 2018)在8000 r·min-1和4 ℃下离心10 min, 用蒸馏水洗涤离心沉淀藻体3次以除去残留培养基.收获的钝顶螺旋藻在无菌条件下接种到模拟废水中, 连续培养7 d, 每天取样检测水质, 每组设置3个平行实验, 结果以平均值和误差表示.为确定钝顶螺旋藻处理海水养殖循环水的最佳接种浓度, 在相同水质条件下, 设置不同微藻接种浓度(0.06~0.21 g·L-1);为研究初始有机碳含量的影响, 固定氮和磷的浓度, 仅改变初始COD(10~100 mg·L-1);探究氮浓度影响时, 固定COD和氮磷比(20 ∶ 1), 改变氮浓度(10~40 mg·L-1)和磷浓度(0.5~2.0 mg·L-1).在实验过程中锥形瓶放置在光照培养箱中, 培养温度为25 ℃, 光暗周期为14 h ∶ 10 h, 光照强度为90 Lx, 每隔8 h摇动烧瓶一次, 防止藻体贴壁.
2.3 反应器启动与运行本研究采用的MPBR反应器实验装置如图 1a所示.MPBR由有机玻璃制成, 内径和有效高度分别为10 cm和38 cm, 有效工作体积为3 L, 反应器A(RA)的膜组件使用膜孔径为0.01 μm的聚偏二氟乙烯(PVDF)膜材料, 有效膜面积为0.2 m2, 反应器B(RB)的膜组件由以PVC板为框架和15 μm无纺布组成, 膜组件的有效过滤面积为0.2 m2.采用9 W的LED灯作外置光源, 安装在反应器外围.反应器底部放置曝气装置, 曝气量为0.3 L·min-1.出水通过膜组件和蠕动泵排出反应器, 反应器底部设置排泥口用来收获微藻.室内温度维持在(25±3) ℃.
 |
| 图 1 反应装置示意图(a)和钝顶螺旋藻的生物干重与光密度的关系(b) Fig. 1 Scliematic diagram of the MPBR reactor (a) and Relationship between biomass of Spirulina plantensis and the optical density (b) |
反应器启动阶段直接将Zarrouk培养基(Zhou et al., 2017)和钝顶螺旋藻接入, 为快速循环掉培养基成分, 加快动态膜的形成, 在启动阶段(运行15 d)持续光照, 设计HRT为12 h.待动态膜形成且出水稳定后, HRT改为48 h, 光暗周期为14 h ∶ 10 h.每30 d收获一次微藻, 即污泥停留时间(SRT)为30 d.
2.4 分析项目与方法微藻生物量(DW, g·L-1)用分光光度法测定(Wang et al., 2020), DW=0.361OD560-0.0637(图 1b).
水样经过0.45 μm醋酸纤维膜过滤, 参照《水和废水的监测分析方法》测定废水中的COD、TN、NH4+-N、NO3--N、NO2--N、TP浓度.采用改良后的Kale法提取微藻中的蛋白质(Cynthia et al., 2010), Folin-酚法(Lowry et al., 1956)测定蛋白质含量, 多糖含量采用苯酚-浓硫酸法测定, 微藻类脂质用甲醇-氯仿法提取(Ge et al., 2016), 叶绿素a用90%的丙酮溶液提取后采用分光光度法测定.
取反应器运行结束后的膜进行梯度脱水处理并烘干喷金后, 进行扫描电镜(JEOL JSM-7600F, 日本)观察(Huang et al., 2018).
2.5 生长情况计算实验过程中微藻比生长速率(Specific Growth Rate, SGR, d-1)、倍增时间(Td, d)、生产速率(PR, mg·L-1·d-1)的计算分别如式(1)~(3)所示(崔伟等, 2019).
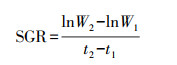
|
(1) |
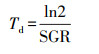
|
(2) |
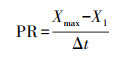
|
(3) |
式中, W1和W2分别为培养开始时t1和t2时刻的微藻生物量;X1为培养开始时的微藻浓度(mg·L-1);Xmax为培养过程中微藻浓度最大值(mg·L-1);Δt为从实验开始至微藻浓度达到最大值所经历的时间间隔(d).
3 结果与讨论(Results and discussion) 3.1 钝顶螺旋藻处理海水养殖循环水的影响因素 3.1.1 污染物去除效能初始碳和氮浓度是导致进水碳氮比波动的直接原因.初始碳、氮浓度和藻接种浓度对污染物去除的影响分别如图 2和表 1所示.可知, 钝顶螺旋藻在研究的接种浓度下对污染物具有良好的处理效果, 接种浓度越大, 污染物去除率越高.硝氮、亚硝氮和总磷的最终去除率均达到100%, 亚硝氮浓度先升高后下降, 总氮去除率达到91%, 表明微藻同化型硝酸盐还原过程中亚硝氮作为中间产物产生, 并继续被还原成氨氮后被钝顶螺旋藻通过GS/GOGAT循环结合到碳骨架中(Perez-garcia et al., 2011), 最终被同化吸收.处理后COD和总氮浓度分别低于20 mg·L-1和1 mg·L-1.当接种浓度为0.12 g·L-1及以上时, 出水总氮浓度低于0.5 mg·L-1, 满足《海水养殖水排放要求》(SC/T 9103—2007)一级标准.
 |
| 图 2 不同藻接种浓度(a~f) 与初始有机碳(g~l)和氮浓度(m~r) 条件下钝顶螺旋藻对COD、TN、NH4+-N、NO2--N、NO3--N及TP的去除效能 Fig. 2 Removal efficiency of Spirulina platensis for COD、TN、NH4+-N、NO2--N、NO3--N and TP in marine aquaculture wastewater with different inoculation volumes(a~f), initial organic carbon(g~l) and nitrogen level(m~r) |
| 表 1 不同藻接种浓度与初始有机碳和氮浓度条件下钝顶螺旋藻对COD、TN、NH4+-N、NO2--N、NO3--N及TP的去除率与去除速率 Table 1 Removal efficiency and removal rate of Spirulina platensis for COD、TN、NH4+-N、NO2--N、NO3--N and TP in marine aquaculture wastewater with different inoculation volumes, initial organic carbon and nitrogen level |
钝顶螺旋藻在研究的有机碳浓度下, 可使出水COD均低于20 mg·L-1, 总磷未检出.增加初始有机碳浓度提高了碳氮比, 进而促进了所有关注污染物的去除.当有机物浓度大于50 mg·L-1, 即碳氮比大于5时, 出水硝氮和亚硝氮未检出.在极低的碳氮比(C/N=1)条件下, 硝氮和总磷被完全去除, 而氨氮、亚硝氮和总氮也实现了较高的去除效率, 分别为87.1%、65.4%和87.7%.这主要归因于钝顶螺旋藻作为一种兼养性原生微生物, 既可以利用培养基中的有机碳异养同化营养物, 也可以通过自养光合作用代谢营养成分(Ge et al., 2016).此外, 碳氮比降低延缓了亚硝氮峰值出现的时间且降低了亚硝氮的累积量, 这主要归因于硝氮与氨氮竞争有机碳;高碳氮比条件下促进了硝氮还原为亚硝氮, 而亚硝氮还原速率小于硝氮造成代谢过程中亚硝氮的累积, 产生的亚硝氮在碳氮比大于3时几乎被完全去除.
钝顶螺旋藻在研究的初始氮浓度下, 对COD的去除效率无显著影响, 出水COD均小于20 mg·L-1.随着初始氮浓度的增加和碳氮比的降低, 氮和磷的去除效率降低, 而去除速率提高, 表明由于碳氮比降低导致异养同化受到抑制.总氮和总磷的去除效率从100%降低到92.2%, 而净去除量从11.4和1.1 mg·L-1提高到29.7和3.7 mg·L-1, 表明低碳氮比条件下光合自养协同异养同化去除污染物.此外, 除初始氮浓度为10 mg·L-1外, 其他氮浓度下亚硝氮持续累积, 可能是由于低碳氮比限制了钝顶螺旋藻的同化型硝酸盐还原过程.
3.1.2 微藻生长状况初始碳、氮浓度和微藻接种浓度对钝顶螺旋藻生长情况及生物质产量的影响如表 2所示.由表可知, 微藻在所探究的接种浓度下均表现出良好的生长状态;藻体蛋白质、多糖和脂质含量无显著差别;随着接种浓度的增加, 比生长速率从0.26 d-1降至0.09 d-1, 而倍增时间从(2.68±0.44) d增加到(7.85±0.61) d.高接种浓度造成微藻细胞之间相互遮挡降低光照强度, 且导致微藻之间对可利用养分的强烈竞争(Liu et al., 2019).因此, 选择合适的接种浓度对于钝顶螺旋藻的培养十分关键.
| 表 2 不同藻接种浓度、初始有机碳及初始氮浓度的海产养殖水中钝顶螺旋藻生长情况及生物质产量 Table 2 Growth and biomass composition of Spirulina platensis in marine aquaculture wastewater with different inoculation volumes, initial organic carbon and nitrogen level |
随着有机碳浓度的增加, 钝顶螺旋藻的生物量和比增长速率分别提高到最大的0.32 g·L-1和0.09 d-1.藻体内蛋白质含量随有机碳浓度和碳氮比提高从566.1 mg·g-1降到488.9 mg·g-1, 而油脂含量由14.9%增到19.3%.Leema等(2010)将海水与Zarrouk培养基混合培养螺旋藻, 收获的微藻脂质含量为8.04%~12.14%, 实验结果表明与完全自养培养相比, 混合培养可以加快微藻的生长速度, 增加脂质积累(Zhan et al., 2017).因此, 在实际应用中可通过增加碳氮比提高生物质和脂质产量.
在研究的初始氮浓度下, 增加氮浓度可提高比生长速率, 倍增时间也随之缩短.在10~30 mg·L-1的氮浓度范围内增加能够提高微藻生物量、蛋白质和多糖含量, 而过高的氮浓度(40 mg·L-1)反而会抑制微藻生长代谢.此外, 提升氮浓度导致碳氮比降低, 藻体油脂含量从16.5%持续下降到14.3%.
综上分析, 在合适的碳和氮浓度范围内, 不论是基于升高有机碳浓度还是降低氮浓度导致的碳氮比提升, 均导致蛋白质含量降低和脂质含量提高.因此, 有机碳、氮、磷浓度和碳氮比共同影响藻体生长.
3.2 膜光合反应器处理海水养殖循环水效能 3.2.1 污染物去除效能图 3展示了两种膜材质的膜光合反应器长期处理性能.在启动期(0~15 d)内, 由于接种过程带入微藻培养基导致出水COD、氮和磷浓度较高, 15 d后培养基被完全替换进入稳定期.稳定期内, 两个反应器均具有良好的碳、氮、磷同步去除性能, 氨氮、亚硝氮、硝氮、总氮和总磷去除效率无显著差别, 分别大于93%、95%、93%、92%和93%, 出水平均总氮和总磷浓度分别满足《海水养殖水排放要求》(SC/T 9103—2007)二级标准和一级标准.两个反应器中COD去除效率差异较为显著, RA出水COD为10.7 mg·L-1, 低于《海水养殖水排放要求》(SC/T 9103—2007)二级标准, 去除率为85.2%;RB出水COD为4.1 mg·L-1, 低于《海水养殖水排放要求》(SC/T 9103—2007)一级标准, 去除率为93.2%.
RA在第1次收获微藻后, 从第2周期开始出水COD、亚硝氮、总氮和总磷浓度出现较大波动, 最高浓度分别为30.0、0.3、1.8和0.4 mg·L-1, 经过20 d反应器自修复, 碳、氮、磷去除效率恢复至平均水平.与马航等(2019)使用青岛大扁藻搭建的微藻膜反应器结果数据相比, 本研究应用钝顶螺旋藻处理海水养殖循环水的过程中不会出现亚硝酸盐的积累.此外, MPBR中对模拟海水养殖循环水的碳、氮、磷处理性能优于批次实验, 可能是由于反应器中生物量相较于批次实验更高.
 |
| 图 3 MPBR系统中COD (a)、TN (b)、NH4+-N (c)、NO2--N (d)、NO3--N (e) 及TP (f) 去除效能 Fig. 3 Removal efficiency of COD(a), TN(b), NH4+-N(c), NO2--N(d), NO3--N(e) and TP(f) in MPBR |
MPBR中微藻生长情况如图 4a所示, RA中微藻持续增长, 生物量在30 d内到最大值0.82 g·L-1, RB在第15 d时微藻结块, 生物量减少, 但第30 d后恢复增长并最终达到0.65 g·L-1(第45 d), 表明系统进行了自我修复.在接下来的两个周期内, RA和RB中微藻生长良好, 最大生物量分别达到0.66~0.71 g·L-1和0.72~0.73 g·L-1.Gao等(2018)使用膜光合生物反应器处理废水时, 稳定期微藻生物量达到1.035~1.524 g·L-1, 这说明膜生物反应器对于微藻具有很好的截留效果, 有助于微藻生物量的积累.
 |
| 图 4 MPBR运行过程中微藻生物量(a) 及其生物质产量(b) Fig. 4 Biomass (a) and content (b) of microalgae in the MPBR |
反应器生物质产量(均以干重计)如图 4b所示.稳定运行阶段RB的多糖平均含量(98.1 mg·g-1)显著高于RA(70.4 mg·g-1), 然而低于文献报道中培养的多糖含量(Peng et al., 2019), 可能是因为海水养殖循环水中有机碳含量较低所致.此外, RA的蛋白质平均含量(622.4 mg·g-1)显著高于RB(544.1 mg·g-1).蛋白质和多糖含量的差异可能归因于多糖更有利于动态生物膜的形成和稳定(Madkour et al., 2012).稳定期RA和RB中油脂含量(16.11%和16.26%)相较于启动期(16.6%和16.65%)略有下降, 其主要原因是启动期高营养的培养基被寡营养的模拟海水养殖循环水替换, 这也验证了批次实验中高碳氮比有利于油脂积累的结论.
启动期内, 随着反应器中各种营养物浓度下降, 作为光合系统I (PSI)的核心光合色素—叶绿素a含量在两个反应器均呈现上升趋势, 微藻通过光合色素含量调控增强自养代谢功能以适应营养物浓度的变化(Villarejo et al., 1995).进入稳定期后叶绿素含量下降, 与生物量变化趋势一致, 表明微藻的光合作用影响并限制了微藻生长(Lorenzen et al., 1967).
3.3 膜光合反应器膜污染情况出水浊度和膜通量是判断动态膜形成的重要参数.由图 5可知, 反应器启动阶段, 由于RA中PVDF膜孔径(0.01 μm)小于微藻粒径, 藻体被阻隔, 出水浊度小于5.0 NTU, 膜通量稳定在7.7~11.5 L·m-2·h-1.RB中无纺布孔径较大导致运行前10 d出水含有部分藻体, 浊度高达97.0 NTU.随着藻体和颗粒物在无纺布表面富集形成生物膜, 出水浊度下降到21.0 NTU并趋于稳定, 膜通量稳定在55 L·m-2·h-1.稳定运行期间(15~105 d), RB的膜通量稳定在55.2 L·m-2·h-1, 无显著降低, 而RA的膜通量由第1周期的17.1 L·m-2·h-1降至第3周期的10.6 L·m-2·h-1, 表明PVDF膜比无纺布动态膜的膜污染更严重.
 |
| 图 5 MPBR反应器出水浊度(a)和膜通量(b) Fig. 5 Turbidity (a) of effluent and membrane flux (b) in the MPBR |
对两种膜组件膜污染前后的形态进行扫描电镜观察.由图 6a~6c可见, 与未使用的PVDF膜相比, 运行105 d后的PVDF膜表面被污染物覆盖, 部分膜孔被堵塞导致膜通量下降.如图 6 d~6f所示, 与未使用的无纺布相比, 无纺布表面分散的滤饼层有助于生物质截留, 部分膜污垢沉积在无纺布的纤维空隙间, 运行过程中RB的膜通量没有显著降低, 表明动态膜膜污染较轻.此外, 无纺布表面的滤饼层经物理清洗后可以去除, 属于可逆污染(程恒卫等, 2015).
 |
| 图 6 MPBR中PVDF (a~c)和无纺布(d~f) 运行前后的SEM图 (a, d: 运行前;b, c, e, f: 运行后) Fig. 6 SEM image of PVDF(a~c) and non-woven fabric(d~f) in MPBR (a, d: before operation; b, c, e, f: after operation) |
1) 本研究确定了钝顶螺旋藻在不同接种浓度、有机碳浓度、氮浓度的海水养殖循环水中的最适运行条件.在原水碳氮比为5时, 钝顶螺旋藻可实现碳、氮、磷高效一体化去除, 出水COD低于20 mg·L-1, 满足《海水养殖水排放要求》(SC/T 9103—2007)一级标准, 总氮和总磷浓度均满足二级标准.添加有机碳或降低氮浓度以提高碳氮比有利于氮、磷的去除.
2) 碳氮比、有机碳、氮和磷浓度共同影响藻体的生长.在合适的碳氮浓度范围内, 碳氮比增加可降低蛋白质含量而提高脂质含量.
3) 钝顶螺旋藻在PVDF膜和无纺布动态膜光合反应器中长期稳定运行期间的总氮、总磷和COD去除效果优于批次实验, 无纺布动态膜反应器的COD去除性能更优.
4) 无纺布动态膜反应器中的多糖含量明显高于PVDF膜反应器, 膜通量也较低, 表明无纺布上形成了稳定的动态生物膜, 膜污染较轻.
Ansari F A, Singh P, Guldhe A, et al. 2017. Microalgal cultivation using aquaculture wastewater: Integrated biomass generation and nutrient remediation[J]. Algal Research, 21: 169-177. DOI:10.1016/j.algal.2016.11.015 |
Anto S, Mukherjee S S, Muthappa R, et al. 2020. Algae as green energy reserve: Technological outlook on biofuel production[J]. Chemosphere, 242: 125079. DOI:10.1016/j.chemosphere.2019.125079 |
Cao L, Wang W, Yang Y, et al. 2007. Environmental impact of aquaculture and countermeasures to aquaculture pollution in China[J]. Environmental Science and Pollution Research, 14(7): 452-462. DOI:10.1065/espr2007.05.426 |
陈凡雨, 徐仲, 尤宏, 等. 2020. 缺氧MBR-MMR处理海水养殖废水性能及膜污染特性[J]. 环境科学, 41(6): 260-268. |
程恒卫, 张彦平, 韩晓栋, 等. 2015. 无纺布动态膜生物反应器处理生活污水的研究[J]. 河北工业大学学报, 44(1): 109-112. |
崔伟, 高峰, 朱凤, 等. 2019. 利用膜-光生物反应器(MPBR)连续培养微藻去除海水养殖废水中营养盐的研究[J]. 浙江海洋学院学报(自然科学版), 38(1): 68-75. DOI:10.3969/j.issn.1008-830X.2019.01.010 |
Cynthia V G, Mdel C G, Francisco G A, et al. 2010. Protein measurements of microalgal and cyanobacterial biomass[J]. Bioresource Technology, 101(19): 7587-7591. DOI:10.1016/j.biortech.2010.04.077 |
Gao F, Peng Y Y, Li C, et al. 2018. Coupled nutrient removal from secondary effluent and algal biomass production in membrane photobioreactor (MPBR): Effect of HRT and longterm operation[J]. Chemical Engineering Journal, 335: 169-175. DOI:10.1016/j.cej.2017.10.151 |
高怡菲, 隗陈征, 李伊晗, 等. 2020. 生物-电氧化法去除海水养殖循环水污染物[J]. 水产学报, 44(7): 1137-1146. |
Ge S, Champagne P. 2016. Nutrient removal, microalgal biomass growth, harvesting and lipid yield in response to centrate wastewater loadings[J]. Water Research, 88: 604-612. DOI:10.1016/j.watres.2015.10.054 |
Huang Z, Wang Y, Jiang L, et al. 2018. Mechanism and performance of a self-flocculating marine bacterium in saline wastewater treatment[J]. Chemical Engineering Journal, 334: 732-740. DOI:10.1016/j.cej.2017.10.076 |
Leema J T, Kirubagaran R, Vinithkumar N V, et al. 2010. High value pigment production from Arthrospira (Spirulina) platensis cultured in seawater[J]. Bioresource Technology, 101(23): 9221-9227. DOI:10.1016/j.biortech.2010.06.120 |
Li J, Zhang H Y, Pei Z G, et al. 2019. Microalgae nourished by mariculture wastewater aids aquaculture self-reliance with desirable biochemical composition[J]. Bioresource Technology, 278: 205-213. DOI:10.1016/j.biortech.2019.01.066 |
刘婷, 蒋丽, 周维芝. 2018. 氮形态对钝顶螺旋藻深度除氮性能的影响[J]. 山东大学学报(工学版), 48(4): 116-121. |
Liu Y, Lv J P, Jia F, et al. 2019. Treatment of real aquaculture wastewater from a fishery utilizing phytoremediation with microalgae[J]. Journal of Chemical Technology & Biotechnology, 94(3): 900-910. |
Lorenzen C J. 1967. Determination of chlorophyll and phaeopigments: Spectrophotometric equations[J]. Limnology and Oceanography, 12(2): 343-346. DOI:10.4319/lo.1967.12.2.0343 |
Lowry O H, Rosebrough N J, Farr A L, et al. 1951. Protein measurement with the Folin phenol reagent[J]. The Journal of Biological Chemistry, 193: 265-275. DOI:10.1016/S0021-9258(19)52451-6 |
Luo G Z, Yoram A, Pan Y F, et al. 2013. Inorganic nitrogen dynamics in sequencing batch reactors using biofloc technology to treat aquaculture sludge[J]. Aquacultural Engineering, 52: 73-79. DOI:10.1016/j.aquaeng.2012.09.003 |
Madkour F F, Kamil A E, Nasr H S. 2012. Production and nutritive value of Spirulina platensis in reduced cost media[J]. The Egyptian Journal of Aquatic Research, 38(1): 51-57. DOI:10.1016/j.ejar.2012.09.003 |
马航, 李之鹏, 柳峰, 等. 2019. 微藻膜反应器处理海水养殖废水性能及膜污染特性[J]. 环境科学, 40(4): 1865-1870. |
Peng Y Y, Gao F, Hang W J, et al. 2019. Effects of organic matters in domestic wastewater on lipid/carbohydrate production and nutrient removal of Chlorella vulgaris cultivated under mixotrophic growth conditions[J]. Journal of Chemical Technology & Biotechnology, 94(11): 3578-3584. |
Perez-garcia O, Escalante F M E, De-bashan L E, et al. 2011. Heterotrophic cultures of microalgae: metabolism and potential products[J]. Water Research, 45(1): 11-36. |
Summerfelt S T. 2003. Ozonation and UV irradiation-an introduction and examples of current applications[J]. Aquacultural Engineering, 28(1/2): 21-36. |
Villarejo A, Orus M I, Martinez F. 1995. Coordination of photosynthetic and respiratory metabolism in Chloreua vulgaris UAM 101 in the light[J]. Physiologia Plantarum, 94(4): 680-686. DOI:10.1111/j.1399-3054.1995.tb00984.x |
Wang Q F, Liu W B, Li X T, et al. 2020. Carbamazepine toxicity and its co-metabolic removal by the cyanobacteria Spirulina platensis[J]. Science of the Total Environment, 706: 135686. DOI:10.1016/j.scitotenv.2019.135686 |
王一笑. 2017. 规模化海水养殖废水处理技术探讨[J]. 农业与技术, 37(10): 245-246. |
姚嘉, 黄天寅, 俞汉青, 等. 2011. 污泥性质对动态膜形成和膜污染的影响[J]. 环境科学学报, 31(8): 1640-1646. |
Zhan J, Rong J, Wang Q. 2017. Mixotrophic cultivation, a preferable microalgae cultivation mode for biomass/bioenergy production, and bioremediation, advances and prospect[J]. International Journal of Hydrogen Energy, 42(12): 8505-8517. DOI:10.1016/j.ijhydene.2016.12.021 |
郑冰冰, 吴怡伟, 李云辉, 等. 2020. 不同碳氮比对海水养殖废水脱氮效果的影响[J]. 环境科学研究, 33(8): 1848-1856. |
Zhou W Z, Li Y T, Gao Y Z, et al. 2017. Nutrients removal and recovery from saline wastewater by Spirulina platensis[J]. Bioresource Technology, 245: 10-17. DOI:10.1016/j.biortech.2017.08.160 |