 2021, Vol. 41
2021, Vol. 41



2. 南京水利科学研究院生态环境研究所, 南京 210029;
3. 河海大学水利水电学院, 南京 210098;
4. 南京市鼓楼区水务局, 南京 210000;
5. 南京市鼓楼区水务设施综合养护中心, 南京 210000
2. Eco-environmental Research Department, Nanjing Hydraulic Research Institute, Nanjing 210029;
3. College of Hydrology and Water Resources, Hohai University, Nanjing 210098;
4. Water Affairs Bureau of Gulou District of Nanjing City, Nanjing 210000;
5. Water Facilities Comprehensive Maintenance Center of Gulou District of Nanjing City, Nanjing 210000
河流是人类生存和可持续发展的基础(马宝珊等, 2021), 城市河流作为城市发展的关键资源和环境载体, 除满足航运、防洪之用, 还具有水源供应、水质净化和景观娱乐等生态功能(罗梅等, 2016; 曹毅等, 2020).随着城市化的快速发展, 大量生活污水和未经处理的工业废水排入水环境容量小、自净能力低的城市河流中, 逐步累积的污染物不仅导致城市出现水体黑臭等环境问题, 而且造成河流富营养化, 破坏城市河流生态系统的平衡(李俊, 2016; Li et al., 2017; 孙玉平等, 2018).因此, 开展城市河流水生态调查研究, 对于城市河流治理与水生态修复具有重要指导意义.
在河流生态系统中, 浮游植物作为重要的初级生产者之一(王苏鹏等, 2019; 夏霆等, 2020; 汪琪等, 2020), 在物质循环、能量流动和维持水生态系统稳定中发挥着重要的作用(陈红等, 2018; Manickam et al., 2020; 代亮亮等, 2020).浮游植物具有结构简单和对环境变化反应灵敏的特点, 其群落结构与分布能够反映出水环境状况的变化规律(Lepistö et al., 2004; Zhao et al., 2005; 吴天浩等, 2019).目前浮游植物群落结构特征分析被广泛应用于河流水环境评价, 已成为城市河流水生态研究的重要内容之一(余海霞等, 2016; 任辉等, 2017; 姜登岭等, 2020).方慷等(2013)从河流水质理化指标和浮游植物物种组成与生物多样性指数等方面, 对大清河水系保定段城市河道进行水体的营养化程度和污染程度评价.马婕等(2020)从浮游植物功能群并结合水文和水质理化指标等方面, 分析了影响苏州城区河道浮游植物功能群变化的主要环境因子.总体而言, 当前关于城市河流水生态的大部分研究主要是分析浮游植物群落结构变化及其与常规水质等环境因子的关系.然而, 为满足城市的防洪需求, 城市河流上往往建设大量的闸泵设施来进行河流水位的调控, 不同频率的水位调控会对不同季节物种的组成有一定影响;此外, 由于河道功能定位不同, 不同河段的城市河流护岸类型在也存在明显差异, 也会影响浮游植物群落结构特征.目前, 关于闸泵水位调节及不同护岸类型对城市河流浮游植物群落结构的影响鲜有报道, 有待进一步深入研究.
基于以上背景, 本文以典型城市闸控河流—江苏省南京市西北护城河为研究区域, 开展不同护岸类型及不同季节的河流水质监测和浮游植物调查, 分析城市河流浮游植物群落结构时空变化特征及其影响因素, 以期为城市河流水生态健康评价和水生态治理提供科学支撑.
2 材料与方法(Materials and methods) 2.1 研究区域概况西北护城河位于江苏省南京市鼓楼区(图 1), 32°4'30"~32°6'0"N, 118°44'30"~118°45'30"E, 外连秦淮河, 全长5.6 km, 汇水面积约为3.95 km2.沿河两岸周边分布着城市公园(小桃园和绣球公园)、绿地、小型湖泊、风景名胜区、密集的城市住宅及商业区域.同时, 由于城市防洪排涝需要, 水务管理部门通过设置闸泵在小桃园和金川门实现不同时期的水位调控(图 1).枯水季节(秋季和冬季), 上游小桃园泵站为保证西北护城河足够的生态流量而保持较为稳定水位(7.3 m)运行, 此时下游水位逐渐抬高, 金川门泵站水位保持在6.5 m上下波动;进入汛期前的平水期(春季), 上游小桃园泵站为满足防洪需要, 会降低水位以腾出足够库容防洪;进入汛期后(夏季), 上下游水位由于闸门水位调控而保持频繁的波动.
 |
| 图 1 南京西北护城河采样点分布和水位变化图 Fig. 1 Distribution of the sampling sites and changes of water level in the Northwest-Moat of Nanjing |
根据西北护城河水环境特征与岸线情况调查, 沿河岸线共布设9个采样点(图 1).其中, S1、S2位于河岸线由硬质衬砌护岸的狭长河道, 宽约8~15 m;S3、S4分布在小桃园公园, 河流水面开阔宽约80~120 m, 闸控水位调节对水体波动影响较弱, 两侧岸线主要为软质护坡构成的生态护岸;S5分布在绣球公园中小型湖泊区域, S6、S7、S8和S9河流两侧护岸均采用硬质条石衬砌而成, 周边主要分布为密集居住区和商业区域, 植被绿化程度较低.采样时间分别在2019年11月(秋季)、2020年2月(冬季)、2020年5月(春季)和2020年8月(夏季).
水体理化指标(酸碱度pH、溶解氧和水温等)采用便携式YSI-6600型多参数检测仪测定, 水样采集参照《地表水和污水检测技术规范》(HJ/T91—2002)进行, 采样前使用河水清洗采水器与水样容器2~3次, 利用采水器汲取水面下0.5 m处水体(何欣霞等, 2019), 装入水样容器中, 每个采样点设置1~2个平行样以避免单次采样的偶然性.采集的水样保存于冰盒, 带回实验室测定, 检查方法遵照《水环境检测规范》(SL219—2013).
采集浮游植物定性样品时, 使用25号筛绢网制作的浮游植物网在采样点水面“∞”字形拖动, 样品收集到50 mL聚乙烯瓶中, 并在现场使用1.0%~1.5%体积样品的鲁哥式液摇匀固定(许志等, 2020).采集定量样品时, 使用采水器在水下0.5 m处采集1 L水样, 加入10 mL鲁哥式液摇匀固定, 带回实验室经分液漏斗静置48 h以上, 浓缩至30 mL摇匀后, 取0.1 mL浓缩液注入0.1 mL浮游植物计数框内, 在10×40倍显微镜鉴定并计数(姚绪姣, 2013).每个样本计数2~3次后取平均值, 浮游植物鉴定及分类参照《中国淡水藻类—系统、分类及生态》, 由于1 mm3的细胞体积接近于1 mg湿重生物量, 故可以直接由藻类的体积换算为生物量(湿重), 单位为mg·L-1(张珊等, 2020; 汪琪等, 2020).
2.3 评价指标与方法浮游植物群落结构特征评价指标选择优势度(Y)、Shannon-Wiener多样性指数(H')和Pielou均匀度指数(J)(李娜等, 2020), 计算公式见式(1)~(3).
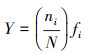
|
(1) |

|
(2) |
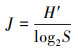
|
(3) |
式中, fi为第i种在所有样品中出现的频率, ni为第i种浮游植物的个体数, N为浮游植物所有种群的总个体数, S为浮游植物中种类的总数.
采用SPSS 23对不同采样点浮游植物群落结构指标进行空间聚类分析, 同时为了使得数据服从正态分布, 对除pH以外的相关性分析数据进行对数处理.采用SPSS 23和R语言corrplot程序包对浮游植物群落结构特征指标(细胞密度、生物量、多样性指数、均匀度指数、硅藻门、绿藻门、蓝藻门及其他门类藻类(包括裸藻门、黄藻门、甲藻门和隐藻门)采用不同门类的生物量)、水质指标(总氮TN、总磷TP、磷酸根PO43-、氮磷比TN∶TP、溶解氧DO)及理化指标(酸碱度pH、水温WT)进行环境因子相关性分析.
3 结果与分析(Results and analysis) 3.1 浮游植物种类组成与优势种不同季节的浮游植物物种组成如图 2所示, 四季共采集到7门90种浮游植物.从物种组成的季节变化(图 2a)来看, 夏季物种数最为丰富(6门55种), 春季(7门42种)和冬季(5门32种)次之, 秋季(6门18种)最少.不同季节浮游植物物种数主要以硅藻门和绿藻门为主, 分别是夏季(硅藻门31种和绿藻门15种)、春季(硅藻门23种和绿藻门10种)、冬季(硅藻门17种和绿藻门8种)及秋季(绿藻门7种和硅藻门5种).从物种组成的空间变化(图 2b)来看, 不同点位的物种数也存在明显的空间差异, 河流在水面宽阔区域(S4、S5)的物种组成与总物种数丰富, 而在岸边带周围属于高密度住宅区(S8、S9)的藻类物种数相对最少.
 |
| 图 2 南京西北护城河浮游植物物种数组成 Fig. 2 Composition of phytoplankton species number in the Northwest-Moat of Nanjing |
表 1为不同季节浮游植物优势种和优势度(Y≥0.02)的统计结果, 4个季节共发现16种优势种, 隶属于7个门类.不同季节优势种存在明显差异, 冬季和春季优势种主要是硅藻门, 夏季优势种主要是硅藻门和绿藻门的小球藻Chlorella vulgaris, 秋季优势种在多个门类均有发现.不同季节中优势度最高的两种藻类是变异直链藻Melosira varians(春季)和黄丝藻Tribonema sp.(秋季), 优势度分别为0.27和0.23.颗粒直链藻Melosira granulata在不同季节中均为优势种, 但其优势度在不同季节存在一定差异.
| 表 1 不同季节浮游植物优势种和优势度 Table 1 Dominant species and dominance of phytoplankton in different seasons |
图 3所示为西北护城河浮游植物的密度和生物量, 四季浮游植物的密度为2.1×103~5.47×106 cells·L-1, 生物量为2×10-3~6.28 mg·L-1.从浮游植物密度来看, 各季节浮游植物门类存在明显的演替规律.夏季浮游植物平均密度最高, 其值为2.52×106 cells·L-1, 但在不同河段浮游植物种类有差别, 比如S3~S5段(生态护岸段)绿藻门浮游植物密度相对较高, 分别占S3、S4、S5浮游植物总密度的81%、94%、74%, 而在硬质护岸的其它河段绿藻门浮游植物密度相对较低, 这是因为生态护岸段水面宽阔、生境较好, 更有利于绿藻门浮游植物生长.自夏入秋, 夏季的绿藻门浮游植物大量消亡, 到秋季则主要以蓝藻门为主.自秋入冬、春季, 由于温度明显较夏季和秋季降低, 蓝藻门浮游植物大量消亡, 到冬季和春季则主要以硅藻门为主.
 |
| 图 3 南京西北护城河四季浮游植物密度(a)与生物量(b) Fig. 3 Cell density (a) and biomass (b) of phytoplankton in the Northwest-Moat of Nanjing |
从浮游植物生物量来看, 不同季节浮游植物门类演替规律与密度有所差异.从浮游植物生物量的季节变化来看, 春、夏季浮游植物平均生物量(1.54 mg·L-1)最大, 其主要原因是温度回升(由冬入春、夏季)更有利于硅藻门浮游植物生长, 硅藻门生物量迅速增加.冬季浮游植物平均生物量最低, 其均值为1.2×10-2 mg·L-1, 这是因为冬季水温不适于浮游植物生长和繁殖, 尤其是蓝藻门浮游植物全河段均未检出.从浮游植物生物量的空间变化来看, 夏季由于S3~S5段(生态护岸段)绿藻门浮游植物大量生长, 因此, 夏季绿藻门生物量占比上升, 而硅藻门生物量占比有所下降.秋季S4~S9段河流中黄藻门浮游植物大量增加, 其均值达到0.27 mg·L-1;虽然秋季蓝藻门平均密度最大, 但由于蓝藻门浮游植物单个细胞质量较小, 因而蓝藻门平均生物量最小.
3.3 浮游植物群落多样性图 4为浮游植物Shannon-Wiener多样性指数和Pielou均匀度指数, 四季多样性指数为0.60~2.35, 均值为1.61, 均匀度指数为0.17~0.59, 均值为0.45.参照浮游植物生物多样性指数评价河流水环境质量标准(方虹, 2019; 管祥洋等, 2020), 多样性指数和均匀度指数指示西北护城水环境质量整体评价结果为中度污染.
 |
| 图 4 南京西北护城河浮游植物多样性指数(a)和均匀度指数(b) Fig. 4 Diversity index (a) and evenness index (b) of phytoplankton in the Northwest-Moat of Nanjing |
从多样性指数来看(图 4a), 夏季多样性指数平均值最高, 其值为1.85, 不同采样点的多样性指数为1.17~2.35, 其中, S4的多样性指数最低, 水质评价处于中度污染.冬季多样性指数平均值最低, 其值为1.49, 不同采样点的指数为0.6~2.08, 由于肘形针尖杆藻Synedra ulna在生物量方面占据主要优势, 从而导致S4的多样性指数最低, 水质评价处于重度污染.
从均匀度指数来看(图 4b), 夏季均匀度指数平均值最高, 其值为0.47, 不同采样点均匀度指数为0.32~0.56, 其中S7处仅检出硅藻门, 并且不同种类的硅藻门分布均匀, 使得S7的均匀度指数最高, 水质评价处于轻度污染.春季均匀度指数均值最低, 其值为0.40, 不同采样点的均匀度指数介于0.25~0.45, S2的均匀度指数最低, 水质评价处于重度污染.
从空间上来看, 多样性指数和均匀度指数在空间各采样点存在明显的差异性.不同采样点存在单一藻类大量繁殖, 从而影响了群落结构的多样性和稳定性.例如, S4在夏季(小球藻)、秋季(黄丝藻)和冬季(肘状针杆藻)的生物量最高, 而且是对应季节的主要优势种, 导致该点在不同季节的多样性指数和均匀度低, 并且S4存在的高生物量和低均匀度特征进一步阐明该区域是典型城市富养化水体.
3.4 浮游植物与环境因子相关性表 2所示为西北护城河环境因子的季节变化, 不同环境因子在不同季节存在差异.水温在季节变化中差异最为明显, 不同季节变化趋势为:夏季>春季>秋季>冬季.总氮呈现秋冬季高、春夏季低的特征, 主要原因是春、夏季闸控水位调节造成水体更新频率变快, 降低了城市河流的总氮含量;而在秋季和冬季, 闸控水位调节的频率降低, 水体更新和流动性降低, 总氮在城市闸控河流内沉积量增加从而导致含量上升.
| 表 2 南京西北护城河环境因子的季节变化 Table 2 Seasonal variations of environmental factors in the Northwest-Moat of Nanjing |
图 5所示为不同季节浮游植物群落指标与环境因子的Pearson相关系数(蓝色代表为正相关, 红色为负相关, 圆圈面积越大且颜色越深表示相关系数越高).水温与浮游植物生物量呈显著正相关(r=0.49, p < 0.01), 表明水温对浮游植物的生长起促进作用.浮游植物生物量与总氮呈显著负相关(r=-0.43, p < 0.01), 表明总氮含量对浮游植物生物量影响较大.不同门类的浮游植物与环境因子相关性结果存在一定差异, 总氮与硅藻门(r=-0.35, p < 0.05)和绿藻门(r=-0.33, p < 0.05)呈显著负相关, 表明总氮对于硅藻门和绿藻门浮游植物的生长影响显著.蓝藻门与氮磷比呈显著正相关(r=0.4, p < 0.05), 高氮磷比对蓝藻门的繁殖影响较大.
 |
| 图 5 浮游植物群落指标与环境因子的Pearson相关系数 Fig. 5 Pearson correlation coefficients of phytoplankton community indicators and environmental factors |
图 6所示为各采样点浮游植物群落结构指标的空间聚类分析结果, 在相似系数10%水平上分为两大群组:Ⅰ类是生态护岸的河流(S3~S5段), Ⅱ类是主要由浆砌条石组成的硬质护岸河段(S1~S2段)和S6~S9段.同时依据空间聚类分析的结果, 统计空间聚类分析的河流浮游植物群落指标, 结果如表 3所示.河流在生态护岸(Ⅰ类)浮游植物密度和生物量明显大于Ⅱ类区域, 但生态护岸(Ⅰ类)浮游植物多样性指数和均匀度指数低于硬质护岸(Ⅱ类).
 |
| 图 6 浮游植物群落指标聚类分析 Fig. 6 Cluster analysis of phytoplankton community indicators |
| 表 3 不同类型的浮游植物群落指标 Table 3 Different types of phytoplankton community indicators |
西北护城河属于典型的城市闸控河流, 水体更新主要依赖于上下游的闸门水位调控, 其对河流生境条件的影响重大.浮游植物种群差异与硅藻门的物种数量变化之间存在很大的关系, 而水流变化对硅藻门具有密切的影响(康元昊等, 2018).通过对西北护城河水位调控、浮游植物物种组成及优势种分布状况的深入分析, 发现西北护城河春、夏两季属于水位频繁调控的汛期, 该时期内硅藻门物种数最多(春季23种和夏季31种), 优势种主要是硅藻门浮游植物;而在水位保持稳定运行的非汛期(秋季), 小桃园和金川门泵站大部分时间均保持稳定水位运行, 此时期内硅藻门物种数最少(秋季5种), 并且仅颗粒直链藻属于优势种.一般来说, 蓝藻门和绿藻门适于在静止或弱紊动水体中生长, 而硅藻门具有较硬的硅质外壳、耐摩擦和耐滚动等特性(张显忠, 2006), 能够在水体流动或紊动环境下获得较大的增殖优势(李飞鹏等, 2017).硅藻门物种数的变化现象进一步阐明了水位调控对硅藻门浮游植物生长影响显著, 主要原因是频繁水位调控(春季和夏季)的水流环境中更适合硅藻门浮游植物的生长.因此, 从城市闸控河流生境长效管理和维持的角度考虑, 可在保障防洪的条件下适当增加水位调节的频率, 有效减少有害藻类(如微囊藻Microcystis sp.)的聚集爆发.
环境因子对浮游植物群落结构具有重要影响, 浮游植物多样性指数是评价浮游植物群落结构的重要指标(Ko et al., 2017; 赵秀侠等; 2020贺玉晓等, 2020).浮游植物多样性评价结果显示, 西北护城河水环境目前处于中度污染状态, 这与不同季节发现的优势藻种均属于中度污染指示物种(如颗粒直链藻、梅尼小环藻Cyclotella meneghiniana、小球藻、黄丝藻、微囊藻和肘状针杆藻)相一致.微囊藻是秋季蓝藻门的一种优势种, 而且会向水体释放微囊藻毒素(许志等, 2020), 在城市河流水环境中应当重点关注.不同的水体环境中影响浮游植物群落结构的环境因子存在一定的差异性(李娜等, 2020; 陈丹丹等, 2020), 温度对浮游植物生长和浮游植物群落分布的影响机理已有大量的研究报道(胡俊等, 2020; 张国庆等, 2020), 即随着温度升高, 部分浮游植物能够更快地进行细胞分裂和生长.西北护城河浮游植物存在一定的演替规律, 冬、春季主要是硅藻门, 随着温度的升高, 绿藻门与蓝藻门分别在夏季和秋季进一步增殖.氮、磷营养盐作为浮游植物生长重要的物质基础, 营养盐的不足或者过量会严重影响浮游植物的生长和群落结构(张珊等, 2020).研究发现, 秋季TN含量较高, 而且TN∶TP高达61.8, 引起蓝藻门的大量繁殖, 同时与蓝藻门和TN∶TP呈显著正相关的结果吻合.
河流护岸类型对城市河流中浮游植物群落结构具有重要影响, 黄奕龙等(2013)通过开展不同类型河道护岸对浮游植物影响的实验发现, 生态护岸为浮游植物的生长和富集提供了良好生境条件.同时, 城市公园区域频繁接受人们在生产、生活和景观娱乐等多方面产生的污染物质.夏季大量降雨径流将冲刷在陆域累积的营养盐进入公园水域, 弱水动力条件(较宽阔的水面)促使大量营养物质沉积.弱水动力条件、汛期低水位和较高的营养盐含量等条件可为绿藻门和蓝藻门在公园区域的水体大量繁殖提供良好的条件, 导致西北护城河S3~S5点位处呈现出较大的浮游植物生物量和密度.未来研究中可以优化布设城市闸控河流的护岸类型, 合理布设生态护岸以减轻人为活动对浮游植物自然生境的影响.
5 结论(Conclusions)1) 西北护城河不同季节浮游植物共发现7门90种, 主要隶属硅藻门和绿藻门, 夏季和春季物种数多, 冬季和秋季物种数少.基于浮游植物生物多样性指数的评价结果表明, 西北护城河水环境总体处于中度污染状态.
2) 西北护城河水位调控对硅藻门浮游植物生长影响显著, 在水流扰动的环境下硅藻门浮游植物可获得更好的增殖优势.从城市闸控河流生境长效管理和维持的角度考虑, 可在保障防洪的条件下适当增加水位调节的频率, 能有效减少有害藻类的聚集爆发.
3) 河流护岸类型对城市河流中浮游植物群落结构具有重要影响, 生态护岸保留了自然河流中的生态循环系统, 可为浮游植物提供更有利的生境条件.
曹毅, 李丽, 陈丽平, 等. 2020. 上海市环城绿带休憩型河道浮游植物群落结构特征[J]. 上海海洋大学学报, 29(5): 734-745. |
陈丹丹, 庞巧珠, 陈晓慧, 等. 2020. 2018年春秋季海南后水湾浮游植物群落结构特征及其与环境因子的关系[J]. 海洋湖沼通报, (5): 113-120. |
陈红, 刘清, 潘建雄, 等. 2019. 灞河城市段浮游生物群落结构时空变化及其与环境因子的关系[J]. 生态学报, 39(1): 173-184. |
代亮亮, 吕敬才, 周维成, 等. 2021. 3种不同营养水平河流浮游植物的群落结构及其与环境因子的相关性研究[J]. 生态学报, 41(2): 62-67. |
方虹. 2019. 九龙江北溪浮游植物群落结构动态及其影响因素[D]. 厦门: 厦门大学
|
方慷, 刘存歧, 杨军. 2013. 大清河水系保定段城市河道浮游植物群落结构及水质评价[J]. 水生态学杂志, 34(3): 25-31. DOI:10.3969/j.issn.1674-3075.2013.03.004 |
管祥洋, 孔明, 张毅敏, 等. 2020. 滆湖水华初期浮游植物群落特征及与水环境因子相关性分析[J]. 环境科学学报, 40(3): 901-914. |
何欣霞, 陈诚, 董建玮, 等. 2019. 江苏里下河腹部地区湖泊湖荡春季浮游植物群落结构和营养状态[J]. 环境科学学报, 39(8): 2626-2634. |
贺玉晓, 刘天慧, 任玉芬, 等. 2020. 北运河秋冬季浮游植物群落结构特征及影响因子分析[J]. 环境科学学报, 40(5): 1710-1721. |
黄奕龙, 朱闻博, 胡仁贵. 2013. 河道护岸结构型式对生态修复影响试验研究[C]. 中国环境科学学会学术年会. 昆明: 5940-5948
|
胡鸿钧, 魏印心. 2006. 中国淡水藻类-系统、分类及生态[M]. 北京: 科学出版社.
|
胡俊, 池仕运, 胡菊香. 2020. 南四湖浮游植群落与环境因子关系的pCCA分析[J]. 环境科学与技术, 43(9): 33-39. |
姜登岭, 赵昊, 邬喜红, 等. 2020. 嘉兴城市河网区高、低水位期浮游植物群落及其与环境因子的典范对应分析[J]. 环境化学, 39(9): 2540-2550. |
Ko C, Lai C, Hsu H, et al. 2017. Decadal phytoplankton dynamics in response to episodic climatic disturbances in a subtropical deep freshwater ecosystem[J]. Water Research, 109: 102-113. DOI:10.1016/j.watres.2016.11.011 |
康元昊, 施军琼, 杨燕君, 等. 2018. 三峡库区汝溪河浮游植物动态及其与水质的关系[J]. 水生态学杂志, 39(6): 23-29. |
Lepistö L, Holopainen A L, Vuoristo H. 2004. Type-specific and indicator taxa of phytoplankton as a quality criterion for assessing the ecological status of Finnish boreal lakes[J]. Limnologica, 34(3): 236-248. DOI:10.1016/S0075-9511(04)80048-3 |
Li J, Li Y, Qian B, et al. 2017. Development and validation of a bacteria-based index of biotic integrity for assessing the ecological status of urban rivers: A case study of Qinhuai River basin in Nanjing, China[J]. Journal of Environmental Management, 196: 161-167. DOI:10.1016/j.jenvman.2017.03.003 |
李飞鹏, 高雅, 张海平, 等. 2015. 流速对浮游藻类生长和种群变化影响的模拟试验[J]. 湖泊科学, 27(1): 44-49. |
李俊. 2016. 武汉典型湖泊浮游植物特征及其影响因子的研究[D]. 武汉: 湖北工业大学
|
李娜, 周绪申, 孙博闻, 等. 2020. 白洋淀浮游植物群落的时空变化及其与环境因子的关系[J]. 湖泊科学, 32(3): 772-783. |
罗梅, 刘昔, 陈国梁, 等. 2016. 城市景观河道浮游植物与水质评价[J]. 环境工程学报, 10(12): 7380-7386. DOI:10.12030/j.cjee.201507237 |
Manickam N, Saravana Bhavan P, Santhanam P, et al. 2020. Phytoplankton biodiversity in the two perennial lakes of Coimbatore, Tamil Nadu, India[J]. Acta ecologica Sinica, 40(1): 81-89. DOI:10.1016/j.chnaes.2019.05.014 |
马宝珊, 徐滨, 魏开金, 等. 2020. 安宁河中游浮游植物群落结构及其与环境因子的关系[J]. 生态学杂志, 39(10): 3332-3341. |
马婕, 申利亚, 何培民, 等. 2021. 苏州城区河道浮游植物功能群演替特征及其对环境因子的响应[J]. 上海海洋大学学报, 30(1): 103-112. |
任辉, 田恬, 杨宇峰, 等. 2017. 珠江口南沙河涌浮游植物群落结构时空变化及其与环境因子的关系[J]. 生态学报, 37(22): 7729-7740. |
孙玉平, 于恒国, 周钦, 等. 2018. 典型富营养化城市河流-浙江温瑞塘河的浮游植物群落类型与季节变化[J]. 湖泊科学, 30(2): 375-384. |
王苏鹏, 陈吉炜, 刘意恒, 等. 2019. 城区河流中沉水植物分布特征及其影响因素分析-以宁波城区内河为例[J]. 湖泊科学, 31(4): 1064-1074. |
汪琪, 黄蔚, 陈开宁, 易齐涛, 等. 2020. 大溪水库浮游植物群落结构特征及营养状态评价[J]. 环境科学学报, 40(4): 1286-1297. |
吴天浩, 刘劲松, 邓建明, 等. 2019. 大型过水性湖泊-洪泽湖浮游植物群落结构及其水质生物评价[J]. 湖泊科学, 31(2): 440-448. |
夏霆, 陈静, 曹方意, 等. 2014. 镇江通江城市河道浮游植物优势种群生态位分析[J]. 长江流域资源与环境, 23(3): 344-350. DOI:10.11870/cjlyzyyhj201403006 |
许志, 陈小华, 沈根祥, 等. 2020. 上海河道浮游植物群落结构时空变化特征及影响因素分析[J]. 环境科学, 41(8): 3621-3628. DOI:10.3969/j.issn.1000-6923.2020.08.043 |
姚绪姣. 2013. 三峡水库香溪河库湾浮游植物群落结构演替规律研究[D]. 宜昌: 三峡大学
|
余海霞, 来勇, 赵佳佳, 等. 2016. 城市河道浮游植物群落多样性指数对水质的指示作用[J]. 化学与生物工程, 33(9): 32-36. DOI:10.3969/j.issn.1672-5425.2016.09.008 |
张国庆, 杨雨玲, 唐爱国, 等. 2020. 新安江流域(屯溪段)浮游植物群落结构及其与环境因子的关系[J]. 生态学杂志, 39(2): 527-540. |
张珊, 商乃萱, 张金羽, 等. 2020. 淮河流域西淝河浮游植物群落结构特征[J]. 水生态学杂志, 41(4): 81-89. |
张显忠. 2006. 三峡库区重庆段泥沙对硅藻生长影响试验研究[D]. 重庆: 重庆大学
|
Zhao M, Lei L, Han B. 2005. Seasonal change in phytoplankton communities in Tangxi reservoir and effecting factors[J]. Journal of Tropical & Subtropical Botany, 13(5): 386-392. |
赵秀侠, 卢文轩, 李静, 等. 2020. 青龙湖冬季浮游植物群落结构及其与环境因子的关系[J]. 水生态学杂志, 41(2): 83-90. |
中国环境监测总站. 2013. HJ/T91-2002地表水和污水检测技术规范[S]. 北京: 中国环境科学出版社
|
中华人民共和国水利部. 2013. SL219-2013水环境检测规范[S]. 北京: 中国水利水电出版社
|